
Last Updated May 28, 2026
Cell theory states that cells are the basic structural and functional units of life, that all organisms are composed of one or more cells, and that new cells arise from preexisting cells. These principles remain foundational because they locate living organization, physiological function, hereditary continuity, development, disease, ecological metabolism, biological repair, and experimental measurement at the cellular level. Cell theory is therefore not merely a historical doctrine from nineteenth-century microscopy. It remains one of the organizing frameworks through which modern biology explains how living systems are built, maintained, reproduced, disrupted, and studied.
This article develops Cell Theory and the Basic Unit of Life as a foundational article within the Biology knowledge series. It treats the cell not only as a visible microscopic compartment, but as a bounded, regulated, metabolically active, information-bearing, experimentally measurable unit of living organization. Cells maintain internal conditions, exchange matter and energy, synthesize biomolecules, express genetic information, divide, differentiate, communicate, respond to stress, and participate in larger systems such as tissues, organs, microbiomes, ecosystems, and evolutionary lineages. To understand the cell is therefore to understand one of the central places where life becomes mechanistically tractable.
Main Library
Publications
Article Map
Biology
Related Topic
Chemistry
Related Topic
Earth Science
Related Topic
Environmental Science
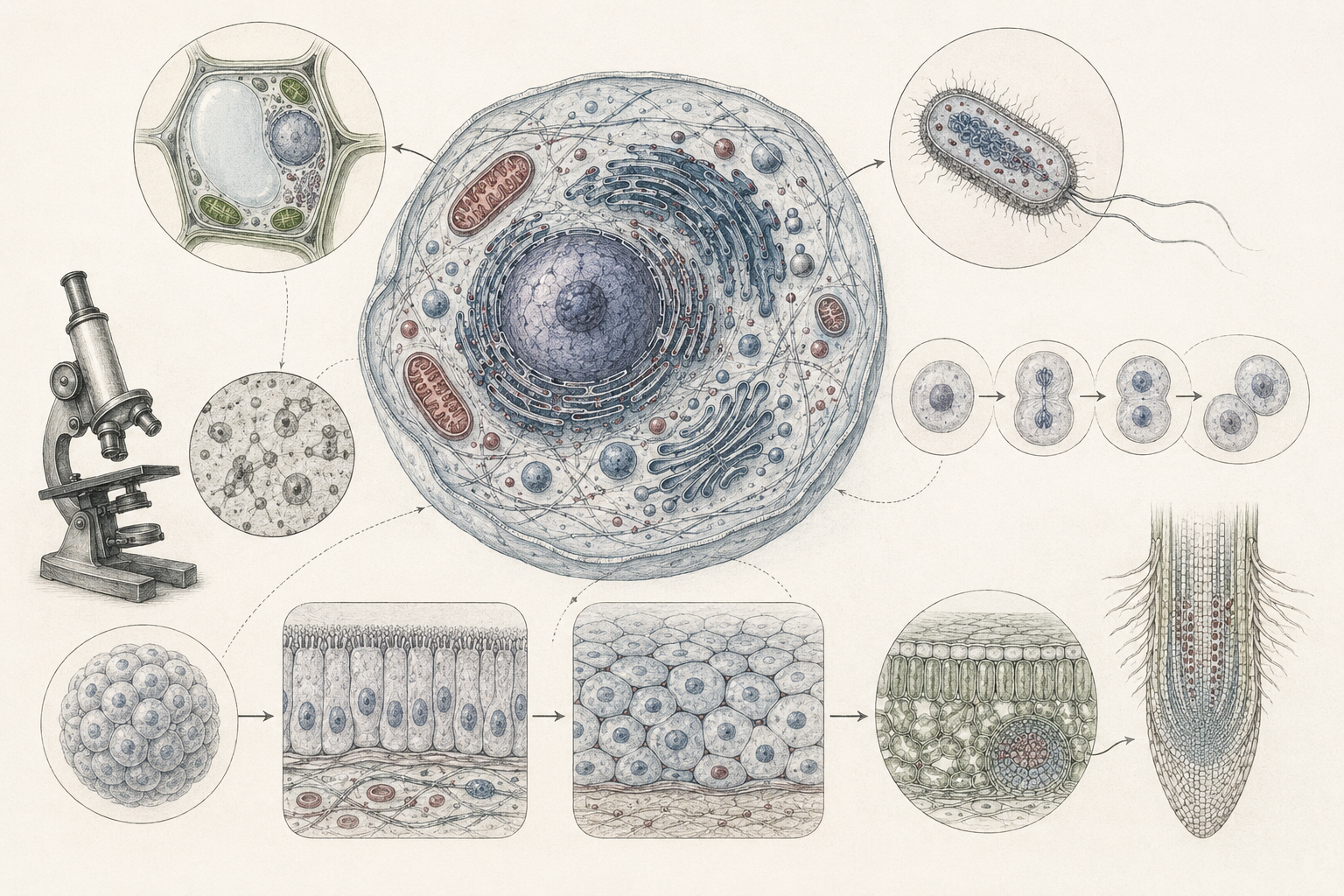
The article develops cell theory across historical microscopy, prokaryotic and eukaryotic organization, membranes, organelles, cell division, cell-cycle regulation, cellular pathology, infection, cancer, immunology, marine biology, microbial ecology, plant science, fungal networks, biotechnology, cell culture, synthetic biology, systems cell biology, single-cell analysis, and computational life science. It shows why cell theory remains indispensable even though modern biology has refined it through molecular biology, genomics, microscopy, biophysics, bioinformatics, and systems modeling.
The article also extends classical cell theory into quantitative and computational biology through exponential and logistic cell growth, doubling time, viability and death-rate estimation, membrane transport, Fickian diffusion, cell-cycle transition models, treatment-response comparisons, cell-density normalization, imaging feature extraction, cell-condition scoring, R workflows, Python workflows, SQL provenance structures, and a linked full-stack GitHub repository containing Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, notebooks, data files, validation notes, and reproducibility documentation.
Core claims of cell theory
Classical cell theory is commonly summarized in three major claims: all living organisms are composed of one or more cells; the cell is the basic unit of structure and function in living systems; and cells arise from preexisting cells. These principles remain foundational because they identify a common organizational level across bacteria, archaea, protists, fungi, plants, animals, and the cellular systems that compose multicellular organisms. Modern biology continues to use the cell as the primary level at which living organization becomes structurally visible, experimentally tractable, and mechanistically explainable.
The first claim, that organisms are composed of cells, gives biology a shared unit beneath the diversity of visible life. A bacterium, a phytoplankton cell, a fungal hypha, a plant leaf, and a human organ may look radically different, but each depends on cellular organization. The second claim, that the cell is the basic unit of structure and function, means that metabolism, transport, signaling, growth, division, gene expression, and response to stress must be explained cellularly even when larger-scale phenomena emerge at the tissue, organismal, population, or ecosystem level. The third claim, that cells arise from preexisting cells, anchors biological continuity in cellular lineage rather than spontaneous generation.
These claims carry strong implications. They mean that physiology, development, heredity, disease, regeneration, immunity, reproduction, and ecological productivity all require cellular explanation. They also prevent biology from treating life as an abstract property detached from organized units. In contemporary science, cell theory is therefore not merely historical background. It remains a framework that supports molecular biology, histology, developmental biology, microbiology, immunology, oncology, regenerative medicine, microbial ecology, marine biology, biotechnology, and computational biology.
Cell theory is powerful because it is both simple and expandable. The core claim that life is cellular remains recognizable, but modern biology has deepened that claim through molecular genetics, membrane biology, organelle function, systems modeling, cell imaging, single-cell sequencing, and computational workflows. The cell is not only a unit seen through the microscope. It is a dynamic system of boundary, information, metabolism, signaling, and continuity.
Historical formation of cell theory
Cell theory emerged from improvements in microscopy and from the broader reorganization of natural history into modern biology. Early microscopists revealed fine tissue structure, but nineteenth-century work by Matthias Schleiden, Theodor Schwann, Robert Remak, Rudolf Virchow, and others helped convert microscopic observation into a general theory of biological organization. The decisive shift was not only that scientists could see cells. It was that cells came to be understood as common units of life, growth, tissue structure, reproduction, and disease.
This transformation changed the scale of biological explanation. Organisms could no longer be understood solely through gross anatomy, external morphology, or visible organs. Growth, tissue formation, pathology, development, and reproduction required cellular explanation. Biology gained a level of analysis that could unify plant and animal life beneath their visible differences. The cell became a common denominator across living diversity.
Cell theory also helped challenge older ideas of spontaneous generation and vitalistic explanations that lacked mechanistic clarity. If new cells arise from preexisting cells, then continuity, division, and lineage become central to the explanation of life. Modern molecular biology later deepened this insight by showing how DNA replication, chromosome segregation, gene expression, and cell-cycle regulation support cellular continuity. In this sense, cell theory was not displaced by modern molecular biology. It was mechanistically enriched by it.
The history of cell theory also illustrates the centrality of instruments and methods in biological science. Microscopes did not merely reveal smaller objects. They changed the conceptual architecture of biology by making the cellular level observable, comparable, and experimentally accessible. Modern imaging, live-cell microscopy, electron microscopy, fluorescence labeling, single-cell sequencing, and computational image analysis continue that same transformation.
Cell theory therefore belongs to the history of both concepts and tools. It shows how a scientific instrument can change what counts as an explanatory unit, how observation can become theory, and how theory can continue to structure research long after its original historical formulation.
Why cells are the basic units of life
The cell is considered the basic unit of life because it is the smallest organized entity capable of maintaining a bounded internal environment, transforming energy, storing and expressing genetic information, synthesizing macromolecules, responding to stimuli, and reproducing through division or participating in a lineage that divides. This definition is functional rather than merely visual. A cell is not simply a microscopic compartment. It is a regulated system in which boundary, information, metabolism, molecular synthesis, and coordinated biochemical activity are coupled.
This point is especially important in multicellular organisms, where cells are highly specialized and often not autonomously viable outside tissue contexts. A neuron, hepatocyte, lymphocyte, epithelial cell, muscle fiber, or plant guard cell performs specialized functions within a larger body. Yet organismal function still depends on cellular processes: membrane transport, intracellular trafficking, mitochondrial energy production, gene regulation, vesicle movement, protein synthesis, cell-cell communication, and controlled proliferation. The organism is a higher-order system, but its core living processes remain cellular in implementation.
Cell theory also matters because it resists both reductionist oversimplification and vague holism. It identifies a minimal level at which life becomes mechanistically tractable while still allowing higher-order organization to remain scientifically meaningful. Cells are not the whole story of life. Tissues, organs, organisms, populations, ecosystems, and lineages matter as real levels of biological explanation. But cells are among the most important places where living order becomes experimentally manageable.
The cellular level is therefore both foundational and relational. A cell is a unit, but not an isolated abstraction. Cells exist in media, tissues, biofilms, symbioses, microbial communities, and ecological environments. They are bounded systems, but their boundaries regulate exchange rather than abolish relation. This is why cell theory remains compatible with systems biology, ecology, and evolutionary biology: it identifies a basic unit without denying the larger systems in which cellular life is embedded.
The cell is also a bridge between chemistry and biology. Biomolecules become living organization only when they are arranged within systems capable of regulation, transport, energy conversion, information processing, and reproduction. Cell theory therefore marks one of the points where molecular matter becomes biologically organized life.
Prokaryotic and eukaryotic organization
Modern cell biology distinguishes two broad cellular plans: prokaryotic cells, which lack a membrane-bound nucleus, and eukaryotic cells, which contain a nucleus and membrane-bound organelles. This distinction is central because it shows that cellular organization is universal across life while also allowing major differences in compartmentalization, genome organization, signaling, metabolism, and intracellular transport. The cell is a shared unit, but cells are not all organized in the same way.
Prokaryotic cells, including bacteria and archaea, are generally smaller and less internally compartmentalized than eukaryotic cells, but they carry out core processes of metabolism, growth, division, signaling, and environmental response with extraordinary efficiency. They dominate many ecological and biogeochemical processes, from nitrogen cycling and decomposition to marine microbial loops and host-associated microbiomes. Their cellular architecture supports rapid growth, metabolic versatility, horizontal gene transfer, and adaptation to diverse environments.
Eukaryotic cells exhibit extensive internal compartmentalization, a dynamic cytoskeleton, endomembrane trafficking, organelle specialization, and complex regulation of gene expression. These features support larger cell size, intracellular transport, multicellularity, specialized tissues, complex development, and compartment-specific biochemical functions. The nucleus, mitochondria, chloroplasts, endoplasmic reticulum, Golgi apparatus, lysosomes, peroxisomes, and cytoskeleton all reflect the eukaryotic expansion of cell theory into a theory of internal organization.
In biomedicine, the prokaryotic-eukaryotic distinction matters because bacterial and human cells differ in ways that are therapeutically exploitable, especially in antimicrobial pharmacology. In ecology, the distinction matters because microbial communities contain cellular lineages with radically different metabolic capacities. In marine biology, it matters because phytoplankton, cyanobacteria, archaea, protists, and animal cells all contribute differently to oceanic productivity and nutrient cycling. In biotechnology, it matters because bacterial, yeast, mammalian, plant, and algal cells each provide different platforms for production, experimentation, and engineering.
This distinction also strengthens rather than weakens cell theory. Biology does not need all cells to be identical for cells to be foundational. The diversity of cellular organization is precisely what makes cell theory scientifically productive: it identifies a shared living unit while opening inquiry into the many ways that unit can be organized.
Membrane compartmentalization and organelle function
Cellular organization depends on selective compartmentalization. The plasma membrane establishes a regulated boundary between intracellular and extracellular environments, controls transport, supports signaling, and preserves ionic and metabolic asymmetry. This boundary is essential because life depends on differences: concentration gradients, electrochemical gradients, pH differences, redox states, solute distributions, and selective exchange. A cell without a regulated boundary cannot maintain the internal conditions required for metabolism, signaling, heredity, and repair.
In eukaryotes, internal membranes further partition function across the nucleus, endoplasmic reticulum, Golgi apparatus, lysosomes, endosomes, mitochondria, chloroplasts, peroxisomes, and other organelles. This partitioning enables spatial separation and coordination of DNA replication, transcription, translation, protein processing, trafficking, degradation, lipid synthesis, detoxification, and energy conversion. Compartmentalization is therefore not merely structural. It is functional architecture.
From a biomedical standpoint, compartmentalization is clinically consequential. Membrane defects contribute to transport disorders, channelopathies, and altered cell signaling. Mitochondrial dysfunction contributes to metabolic, neuromuscular, cardiovascular, and neurodegenerative disease. Lysosomal dysfunction underlies storage disorders. Abnormal trafficking, organelle stress, signaling dysfunction, or protein-processing failure contributes to cancer, immune dysregulation, infectious disease, metabolic syndromes, and inherited cellular disorders.
The same logic extends into ecology and marine biology. Osmoregulation, ion transport, intracellular signaling, photosynthesis in phytoplankton, stress response under salinity or hypoxia, symbiotic cellular exchange, and adaptation to temperature or pH change all depend on membrane and organelle function. Cellular compartmentalization is therefore a basis not only for biomedical explanation but also for environmental and ecological physiology.
Compartmentalization is one of the reasons the cell is more than a container. It is a structured interior in which biochemical processes are localized, sequenced, protected, and coordinated. Cell theory becomes much deeper once the cell is understood as an internally organized system rather than a simple microscopic unit.
Cell division, the cell cycle, and biological continuity
The claim that all cells arise from preexisting cells links cell theory directly to cell division and the cell cycle. Mitosis underlies growth, tissue renewal, development, wound repair, immune expansion, and replacement of damaged or short-lived cells. Meiosis underlies gamete formation, genetic recombination, and intergenerational continuity in sexually reproducing organisms. Binary fission supports prokaryotic proliferation. Across these forms, cellular continuity depends on regulated reproduction of cellular organization.
Modern cell biology adds mechanistic specificity by describing the cell cycle as a regulated sequence controlled by checkpoints, cyclin-dependent kinases, DNA replication controls, spindle-attachment surveillance, damage-response pathways, and programmed decisions between proliferation, arrest, differentiation, senescence, and death. Cell division is not merely a doubling event. It is a highly regulated process that must preserve genomic integrity, coordinate cytoplasmic and organelle inheritance, and respond to environmental and internal signals.
This regulatory architecture is central to biomedical science. Excess proliferation, failed checkpoint control, defective apoptosis, genomic instability, or disrupted signaling contributes to cancer. Insufficient proliferation or impaired renewal contributes to developmental defects, marrow failure, immune deficiency, degenerative processes, infertility, and poor wound healing. Cell theory therefore does not end with the statement that cells divide. It includes the mechanistic logic by which division is licensed, executed, arrested, repaired, or terminated under physiological and pathological conditions.
Cell division is equally important in ecology and marine biology, where bloom dynamics, microbial turnover, plankton growth, larval development, tissue regeneration, and growth under environmental stress all depend on proliferation and survival at the cellular level. Biological continuity may be observed at organismal, population, and lineage scales, but it is executed cellularly.
Cell division also connects cell theory to heredity. A cell must not only reproduce its boundary and cytoplasm; it must transmit genetic information and restore functional organization in daughter cells. Cellular continuity is therefore both material and informational.
Cell theory in pathology and biomedicine
Biomedical science depends on cell theory because pathology is often cellular before it is systemic. Histology, pathology, oncology, microbiology, immunology, regenerative medicine, toxicology, and pharmacology all require understanding how cells are structured, how they communicate, how they respond to injury, and how they fail. Diagnostic microscopy, tissue biopsy interpretation, cytology, immunohistochemistry, molecular pathology, and flow cytometry all assume that disease can be localized to altered cellular morphology, proliferation, differentiation, signaling, metabolism, or death.
This cellular perspective is mechanistic as well as descriptive. Infection depends on host-pathogen interaction at cellular interfaces. Cancer depends on altered proliferation, genomic instability, immune evasion, and dysregulated signaling. Immune responses depend on cellular lineage specification, receptor signaling, trafficking, cytokine production, and effector function. Neurodegenerative disorders often involve protein aggregation, mitochondrial stress, glial response, and neuronal vulnerability. Metabolic disease involves altered insulin signaling, mitochondrial function, lipid handling, and cellular energy balance.
Drug discovery frequently begins by asking how a compound alters receptors, enzymes, ion channels, signaling pathways, organelles, gene expression, cell viability, or phenotype in defined cell populations. Cell culture, organoids, induced pluripotent stem cells, engineered tissues, and high-content screening all extend cell theory into experimental platforms. Medical relevance therefore strengthens rather than narrows cell theory. It shows that the cell is not only a conceptual unit of life but also an operational unit of diagnosis, intervention, and biomedical explanation.
Cell theory also helps medicine avoid purely organ-level reduction. A damaged organ is not only a damaged structure. It is an altered cellular environment involving signaling, inflammation, metabolism, vascular supply, immune response, fibrosis, regeneration, and cell death. Clinical disease becomes intelligible when cellular processes are connected to tissue and organismal consequences.
Infection, immunity, and cellular boundaries
Cell theory is central to understanding infection because pathogens must interact with cells. Viruses enter host cells and depend on cellular machinery for replication. Bacteria adhere to surfaces, invade tissues, or interact with immune cells. Fungi and parasites often alter cellular environments, immune responses, or tissue integrity. Host defense depends on cells that recognize, signal, migrate, kill, repair, and remember. Infection is therefore not only an organism-level event. It is a cellular and molecular encounter.
Immunity is similarly cellular. Innate immune cells recognize danger, phagocytose pathogens, release inflammatory signals, and recruit other cells. Adaptive immune cells proliferate, differentiate, form memory populations, and execute highly specific responses. Antigen presentation, cytokine signaling, antibody production, T-cell activation, immune tolerance, and inflammation all depend on cellular organization and communication. The immune system is a distributed tissue-level and organism-level system, but it operates through cells.
This cellular view also matters for disease ecology. Pathogen transmission, host susceptibility, vector competence, environmental survival, and immune response all have cellular dimensions. Climate stress, nutrition, pollution, microbiome disruption, and habitat change can alter cellular physiology and immunity, reshaping disease outcomes. Cell theory therefore connects infection biology to ecology, public health, and environmental change.
Cellular boundaries are especially important here. A membrane is not merely a wall. It is a regulated interface through which recognition, entry, exclusion, signaling, transport, and immune response occur. Infection and immunity both depend on the biology of cellular boundaries.
Ecological relevance of cell theory
Cell theory matters to ecology because ecological systems are built from organisms whose capacities are grounded in cells. Primary production, decomposition, nutrient cycling, symbiosis, stress tolerance, reproduction, and population persistence all depend on cellular metabolism, transport, signaling, and division. Ecology may often be studied at the level of populations, communities, and ecosystems, but those larger processes remain anchored in the physiology and viability of cells.
Microbial ecology makes this especially clear. Bacteria, archaea, fungi, protists, and microalgae drive decomposition, nutrient turnover, disease dynamics, methane production, nitrogen cycling, sulfur cycling, carbon transformation, and biogeochemical feedbacks. These ecological functions depend on cellular organization: membranes, enzymes, genes, transporters, metabolic pathways, and regulatory systems. A soil, wetland, lake, reef, or ocean cannot be understood ecologically without understanding cellular activity at microscopic scales.
Environmental disturbance also often acts through cellular mechanisms. Temperature stress alters proteins, membranes, and metabolism. Salinity changes affect osmotic balance. Hypoxia affects energy production. Pollutants alter membranes, enzymes, gene expression, and cell viability. Acidification affects ion regulation and biomineralization. Drought changes plant cell water potential and microbial activity. Cell theory therefore provides a bridge between environmental stress and ecosystem-scale consequence.
This bridge is increasingly important in sustainability science. Climate change, pollution, habitat degradation, eutrophication, and ocean acidification become biologically consequential because they change the conditions under which cells maintain metabolism, membranes, growth, signaling, and reproduction. Ecosystem disruption often begins with cellular stress.
Marine biology relevance of cell theory
Marine biology gives cell theory a distinctive environmental frame. Many marine processes depend on cellular physiology under conditions of salinity, pressure, temperature variation, nutrient limitation, light gradients, changing oxygen availability, and shifting pH regimes. Phytoplankton productivity, microbial loops, coral symbiosis, larval development, osmoregulation, biomineralization, and stress responses to warming or acidification are all cellular before they become organismal or ecosystem-level phenomena.
Marine systems also reveal how strongly cellular life is conditioned by environmental gradients. Membrane transport, intracellular ion balance, photosynthetic machinery, mitochondrial function, nitrogen metabolism, carbon concentrating mechanisms, and signaling pathways all respond to marine conditions in ways that matter for population dynamics and ecosystem resilience. A phytoplankton bloom, for example, may appear as a large-scale oceanographic event, but it depends on cellular growth, nutrient uptake, photosynthesis, viral lysis, grazing, and mortality.
Coral reefs illustrate the same principle. Coral bleaching involves breakdown in cellular and symbiotic relationships between coral hosts and photosynthetic dinoflagellates. Ocean acidification affects cellular and molecular processes involved in calcification and ion regulation. Hypoxia affects energy metabolism and survival. For marine biologists, cell theory is therefore not only a background principle. It is a way of locating ocean-scale processes in living units that can be studied mechanistically.
This is also why marine biotechnology and ocean monitoring increasingly depend on cellular and molecular methods: flow cytometry, environmental DNA, microscopy, metagenomics, pigment analysis, stress biomarkers, and single-cell approaches. Ocean biology is not less cellular because it is large-scale. Its scale makes cellular mechanisms even more important.
Plant, soil, microbial, and fungal relevance
Plant biology depends deeply on cell theory. Plant cells are organized through cell walls, vacuoles, chloroplasts, plasmodesmata, membranes, cytoskeletons, and specialized tissues. Photosynthesis, water transport, stomatal regulation, growth, defense, reproduction, and development all require cellular explanation. Plant form emerges through coordinated cell division, expansion, differentiation, and signaling. Even whole-plant responses to drought, heat, salinity, pathogens, and nutrient limitation are mediated through cellular processes.
Soil biology and fungal biology also show the power of cell theory. Soil microbial cells transform organic matter, cycle nutrients, fix nitrogen, respire carbon, produce extracellular enzymes, and shape soil structure. Fungal hyphae extend cellular organization across networks that absorb nutrients, decompose matter, and form symbioses with plants. These systems complicate simple ideas of cellular individuality because fungal filaments, microbial biofilms, and plant plasmodesmata create interconnected living networks. Yet they do not overturn cell theory. They refine it by showing how cells can be bounded and relational at the same time.
Biofilms are especially important. Bacterial and fungal cells in biofilms behave differently from isolated planktonic cells because extracellular matrices, gradients, signaling, and community structure change cellular physiology. This matters in medicine, environmental engineering, agriculture, wastewater treatment, marine fouling, and microbial ecology. Cell theory remains foundational, but modern biology increasingly studies cells in collective and environmental contexts.
These examples show why cell theory is not restricted to animal cells or medical laboratories. It is a general framework for understanding living organization across plants, fungi, soils, microbes, marine systems, and ecosystems.
Biotechnology and computational relevance
Biotechnology extends cell theory into applied systems of intervention, production, and analysis. Cell culture, microbial engineering, fermentation, assay development, gene delivery, synthetic biology, vaccine production, drug screening, biosensing, high-content imaging, and regenerative medicine all depend on the cell as a manipulable, measurable unit. Once cellular behavior becomes experimentally tractable, it becomes possible to design workflows around growth, signaling, viability, differentiation, metabolism, gene expression, and stress response.
Computational methods deepen this transition. Image analysis, growth modeling, single-cell RNA sequencing, cytometry, transcriptomic workflows, proteomics, metabolic modeling, and simulation-based hypothesis testing all treat the cell not only as a biological unit but also as a source of structured data. A cell can be counted, imaged, segmented, classified, modeled, perturbed, and linked to molecular data. In that sense, modern cell theory is also a data architecture.
This computational turn matters because biological interpretation increasingly depends on reproducibility. Cell counts need time stamps, treatment conditions, plate layouts, passage numbers, culture media, imaging metadata, normalization methods, and uncertainty estimates. Single-cell workflows need provenance, preprocessing records, quality control, feature extraction, and transparent assumptions. Cell theory has become not only conceptual and experimental, but computational and infrastructural.
Biotechnology also raises the stakes of cellular interpretation. Engineered cells, organoids, cell therapies, microbial production systems, and synthetic circuits all depend on the assumption that cellular behavior can be characterized, perturbed, controlled, and measured. The more powerful the intervention, the more important it becomes to understand the cell as a dynamic living system rather than as a passive container for genetic instructions.
Mathematical lens
Modern cell biology is not only descriptive and mechanistic. It is also quantitative. Cell populations grow over time, molecules diffuse across membranes, signaling pathways amplify or dampen responses, treatments alter survival and proliferation, and imaging workflows quantify cellular morphology. Cell theory therefore opens into mathematical, statistical, and computational analysis of how cells behave under defined conditions.
If a population of cells grows under unconstrained conditions, early growth can often be approximated by an exponential model:
Interpretation: Exponential growth describes cell-population increase when per-capita growth rate remains approximately constant.
where \(N(t)\) is the number of cells at time \(t\), \(N_0\) is the initial number of cells, and \(r\) is the per-capita growth rate. From this expression, the doubling time is:
Interpretation: Doubling time converts growth rate into the time required for a cell population to double.
This model is useful in cell culture, microbiology, tumor biology, pharmacology, marine microbial growth, and ecological physiology.
If growth is limited by space, nutrients, waste accumulation, contact inhibition, immune response, or treatment, a logistic model may be more appropriate:
Interpretation: Logistic growth distinguishes early growth potential from density-limited or resource-limited cellular expansion.
where \(K\) is carrying capacity.
Transport across membranes and tissues can often be approximated through diffusion:
Interpretation: Fickian diffusion relates flux to the concentration gradient across distance.
where \(J\) is flux, \(D\) is the diffusion coefficient, and \(\frac{dC}{dx}\) is the concentration gradient. This relation is relevant to nutrient uptake, oxygen delivery, intracellular gradients, signaling-molecule movement, salinity stress, drug exposure, and transport-limited assays.
Under stress, treatment, starvation, toxic exposure, or unfavorable culture conditions, viable cell abundance may decline approximately as:
Interpretation: Viability decay models the loss of viable cells through time under stress or treatment.
where \(L(t)\) is viable cell abundance at time \(t\), \(L_0\) is initial viable abundance, and \(k\) is a loss or death-rate parameter.
A simple compartment model for cell-cycle transition can be written as:
Interpretation: A compartment model represents regulated movement of cells through simplified cell-cycle states.
where \(G_1\), \(S\), and \(G_{2M}\) represent cell-cycle compartments, while \(k_1\), \(k_2\), and \(k_M\) represent transition rates. This is a simplified model, but it illustrates how cellular continuity can be represented as regulated movement across states.
Variables, units, and cellular interpretation
Quantitative cell biology depends on variables that connect cell abundance, growth, viability, transport, and cell-cycle state to biological interpretation. The table below summarizes several central quantities.
| Symbol or Term | Meaning | Typical Unit or Scale | Cellular Interpretation |
|---|---|---|---|
| \(N(t)\) | Cell abundance at time \(t\) | cells, colony-forming units, nuclei, reads, or biomass proxy | Measured or modeled number of cells in a population |
| \(N_0\) | Initial cell abundance | same as \(N(t)\) | Starting cell number for growth analysis |
| \(r\) | Per-capita growth rate | per unit time | Rate of cellular population increase under specified conditions |
| \(t_d\) | Doubling time | time | Time required for cell abundance to double |
| \(K\) | Carrying capacity | same as \(N(t)\) | Approximate upper cell abundance supported by limiting conditions |
| \(J\) | Flux | amount per area per time | Rate of transport across a membrane, tissue, or spatial gradient |
| \(D\) | Diffusion coefficient | area per time | How rapidly a molecule spreads through a medium |
| \(C\) | Concentration | molarity, mass per volume, or assay unit | Amount of solute, drug, nutrient, or signaling molecule per volume |
| \(x\) | Distance | length | Spatial dimension over which concentration changes |
| \(L(t)\) | Viable cell abundance | viable cells, colony-forming units, or viability proxy | Remaining viable cells after stress or treatment |
| \(k\) | Viability-loss or death-rate parameter | per unit time | Rate at which viable cells are lost under a condition |
| \(G_1, S, G_{2M}\) | Cell-cycle compartments | fractions, proportions, or cell counts | Cells distributed across simplified cell-cycle states |
| \(k_1, k_2, k_M\) | Cell-cycle transition rates | per unit time | Rates of movement between cell-cycle compartments |
The table shows why cell theory becomes more powerful when paired with measurement. A cell can be counted, timed, perturbed, imaged, classified, and modeled. But each quantity must be interpreted in relation to the biological system, measurement method, and experimental context.
Worked example: growth rate and doubling time
Suppose a culture begins with \(N_0=1.0\times10^5\) cells and grows to \(4.0\times10^5\) cells in 48 hours during its early exponential phase. Under the exponential growth model:
Interpretation: Cell abundance changes according to initial abundance, growth rate, and elapsed time.
Substituting the observed values gives:
Interpretation: The observed cell-count change can be used to estimate the growth rate.
Dividing both sides by \(1.0\times10^5\):
Interpretation: The culture increased fourfold during the observation interval.
Taking the natural logarithm:
Interpretation: The estimated growth rate is approximately 0.0289 per hour.
The doubling time is:
Interpretation: At this estimated rate, the cell population doubles approximately every 24 hours.
This simple calculation turns raw cell-count data into an interpretable biological parameter. Similar calculations are routine in cell culture, microbial growth experiments, tumor kinetics, plankton growth studies, and treatment-response work. The result is not a complete explanation of cellular behavior, but it provides a transparent baseline for comparing conditions.
Computational modeling
Computational modeling helps make cell theory operational because cellular behavior is dynamic, measurable, and often condition-dependent. Cell populations grow, stop, die, differentiate, migrate, signal, and respond to perturbation. Growth curves can estimate rate and doubling time. Viability models can quantify treatment response. Diffusion models can estimate transport constraints. Cell-cycle models can formalize transitions across cellular states. Imaging pipelines can extract size, shape, intensity, texture, and spatial features from large numbers of cells.
The selected examples below focus on compact, reusable workflows: exponential growth fitting, viability-loss estimation, cell-condition scoring, logistic growth simulation, membrane flux calculation, and cell-cycle compartment modeling. The GitHub repository can extend the same logic into richer workflows for high-content imaging, cell-density normalization, treatment-response comparisons, SQL provenance, multi-language numerical examples, notebooks, and validation scripts.
The purpose is not to reduce the cell to computation. The purpose is to make cellular evidence inspectable. A cell-biology claim becomes stronger when the measurement, model, parameter, assumption, and biological interpretation are documented together.
R workflow: cell growth, viability, and condition scoring
R is especially useful for cell biology because it supports statistical modeling, experimental comparison, and reproducible tabular reporting. The following workflow estimates exponential cell growth, estimates treatment-associated viability loss, and applies a transparent cell-condition scoring scaffold.
# Cell Growth, Viability, and Condition Scoring
#
# This workflow demonstrates three quantitative cell-biology tasks:
#
# 1. Estimate exponential growth rate and doubling time.
# 2. Estimate viability loss under treatment or stress.
# 3. Create a transparent cell-condition score.
#
# These examples can be adapted for cell culture, microbial growth,
# marine phytoplankton studies, cytotoxicity assays, drug screening,
# tissue models, or environmental-stress experiments.
library(tibble)
library(dplyr)
# ------------------------------------------------------------
# 1. Estimate exponential growth rate from cell-count data
# ------------------------------------------------------------
growth_data <- tibble(
time_h = c(0, 12, 24, 36, 48),
cells = c(1.0e5, 1.4e5, 2.0e5, 2.8e5, 4.0e5)
)
growth_model <- lm(log(cells) ~ time_h, data = growth_data)
growth_summary <- tibble(
growth_rate_per_h = coef(growth_model)[["time_h"]],
estimated_initial_count = exp(coef(growth_model)[["(Intercept)"]]),
doubling_time_h = log(2) / growth_rate_per_h,
r_squared_log_space = summary(growth_model)$r.squared
)
growth_fit <- growth_data %>%
mutate(
predicted_cells = exp(predict(growth_model)),
residual_log_scale = resid(growth_model)
)
# ------------------------------------------------------------
# 2. Estimate viability loss under treatment conditions
# ------------------------------------------------------------
viability_data <- tibble(
time_h = c(0, 12, 24, 36, 48),
viable_cells = c(1.0e6, 8.2e5, 6.3e5, 4.6e5, 3.2e5)
)
viability_model <- lm(log(viable_cells) ~ time_h, data = viability_data)
loss_rate <- -coef(viability_model)[["time_h"]]
viability_summary <- tibble(
loss_rate_per_h = loss_rate,
half_life_h = log(2) / loss_rate,
r_squared_log_space = summary(viability_model)$r.squared
)
viability_fit <- viability_data %>%
mutate(
predicted_viable_cells = exp(predict(viability_model)),
residual_log_scale = resid(viability_model)
)
# ------------------------------------------------------------
# 3. Transparent heuristic cell-condition scoring
# ------------------------------------------------------------
cell_conditions <- tibble(
condition = c("control", "nutrient_limited", "hypoxic", "drug_treated"),
membrane_integrity = c(0.92, 0.78, 0.70, 0.62),
metabolic_activity = c(0.88, 0.55, 0.46, 0.40),
proliferation_capacity = c(0.84, 0.48, 0.42, 0.30),
genomic_stability = c(0.90, 0.82, 0.76, 0.68),
stress_penalty = c(0.12, 0.42, 0.55, 0.68)
) %>%
mutate(
cell_condition_score =
0.22 * membrane_integrity +
0.24 * metabolic_activity +
0.20 * proliferation_capacity +
0.20 * genomic_stability +
0.14 * (1 - stress_penalty),
condition_class = case_when(
cell_condition_score >= 0.75 ~ "strong_cell_condition",
cell_condition_score >= 0.50 ~ "moderate_cell_condition",
TRUE ~ "constrained_cell_condition"
)
)
print(round(growth_summary, 5))
print(round(growth_fit, 3))
print(round(viability_summary, 5))
print(round(viability_fit, 3))
print(cell_conditions %>% mutate(across(where(is.numeric), ~ round(.x, 3))))This R workflow is useful because it yields directly interpretable quantities: an estimated growth rate, an estimated doubling time, a treatment-associated viability half-life, and an explicit condition score. A working scientist could extend it to treatment comparisons, confidence intervals, mixed models, nonlinear logistic fits, imaging features, or replicate-aware experimental designs.
Python workflow: logistic growth, membrane flux, and cell-cycle states
Python is useful for simulation, numerical modeling, pipeline design, and reproducible computational cell biology. The following workflow simulates logistic growth under different conditions, estimates membrane flux under concentration gradients, and models simplified cell-cycle compartment transitions.
"""
Computational Cell Theory Workflow
This workflow demonstrates three quantitative cell-biology tasks:
1. Simulate logistic cell growth under different experimental conditions.
2. Estimate membrane flux under concentration gradients using Fick's law.
3. Simulate simplified cell-cycle compartment transitions.
The examples are compact, but the same structures can be extended to
cell culture, microbial growth, marine phytoplankton studies, treatment
response, drug exposure, tissue diffusion, single-cell workflows,
and systems cell biology.
"""
from __future__ import annotations
import numpy as np
import pandas as pd
def logistic_growth(
time: np.ndarray,
initial_count: float,
growth_rate: float,
carrying_capacity: float,
) -> np.ndarray:
"""
Calculate logistic cell-growth trajectory.
"""
if initial_count <= 0:
raise ValueError("Initial cell count must be positive.")
if carrying_capacity <= 0:
raise ValueError("Carrying capacity must be positive.")
if initial_count > carrying_capacity:
raise ValueError("Initial count should not exceed carrying capacity.")
return carrying_capacity / (
1.0
+ ((carrying_capacity - initial_count) / initial_count)
* np.exp(-growth_rate * time)
)
def compare_growth_conditions() -> pd.DataFrame:
"""
Simulate logistic cell growth under multiple conditions.
"""
time_h = np.linspace(0, 96, 400)
initial_count = 1.0e5
carrying_capacity = 1.0e6
conditions = {
"control": 0.035,
"treated": 0.020,
"nutrient_limited": 0.015,
}
rows = []
for condition, growth_rate in conditions.items():
trajectory = logistic_growth(
time=time_h,
initial_count=initial_count,
growth_rate=growth_rate,
carrying_capacity=carrying_capacity,
)
rows.append(
pd.DataFrame(
{
"condition": condition,
"time_h": time_h,
"cell_count": trajectory,
"growth_rate_per_h": growth_rate,
"doubling_time_h_initial": np.log(2.0) / growth_rate,
}
)
)
return pd.concat(rows, ignore_index=True)
def membrane_flux_example() -> pd.DataFrame:
"""
Calculate flux under concentration gradients using Fick's law.
J = -D dC/dx
"""
diffusion_coefficient = 2.0e-6 # cm^2/s
distance_cm = np.linspace(0.001, 0.02, 20)
concentration_inside = 1.0
concentration_outside = 0.2
gradient = (concentration_outside - concentration_inside) / distance_cm
flux = -diffusion_coefficient * gradient
return pd.DataFrame(
{
"distance_cm": distance_cm,
"concentration_gradient": gradient,
"flux_units": flux,
}
)
def simulate_cell_cycle(
t_max: float = 48.0,
dt: float = 0.01,
k1: float = 0.10,
k2: float = 0.08,
km: float = 0.06,
) -> pd.DataFrame:
"""
Simulate simplified cell-cycle compartment transitions.
"""
time_h = np.arange(0, t_max + dt, dt)
g1 = np.zeros_like(time_h)
s_phase = np.zeros_like(time_h)
g2m = np.zeros_like(time_h)
g1[0] = 0.70
s_phase[0] = 0.20
g2m[0] = 0.10
for i in range(1, len(time_h)):
dg1 = 2.0 * km * g2m[i - 1] - k1 * g1[i - 1]
ds = k1 * g1[i - 1] - k2 * s_phase[i - 1]
dg2m = k2 * s_phase[i - 1] - km * g2m[i - 1]
g1[i] = max(g1[i - 1] + dg1 * dt, 0.0)
s_phase[i] = max(s_phase[i - 1] + ds * dt, 0.0)
g2m[i] = max(g2m[i - 1] + dg2m * dt, 0.0)
return pd.DataFrame(
{
"time_h": time_h,
"G1_fraction": g1,
"S_fraction": s_phase,
"G2M_fraction": g2m,
"total_fraction": g1 + s_phase + g2m,
}
)
def main() -> None:
"""
Run compact cell-theory computational examples.
"""
growth_df = compare_growth_conditions()
flux_df = membrane_flux_example()
cell_cycle_df = simulate_cell_cycle()
print("Logistic growth comparison:")
print(growth_df.head(12).round(3).to_string(index=False))
print(growth_df.tail(12).round(3).to_string(index=False))
print("\nMembrane flux under concentration gradients:")
print(flux_df.round(8).to_string(index=False))
print("\nSimplified cell-cycle compartment model:")
print(cell_cycle_df.head(12).round(4).to_string(index=False))
print(cell_cycle_df.tail(12).round(4).to_string(index=False))
if __name__ == "__main__":
main()This Python workflow is useful because it converts cell theory into analyzable behavior. Cells are not only structures; they grow under constraints, transport molecules across boundaries, and move through regulated states. These models are simplified, but they provide transparent baselines for experimental comparison.
GitHub repository
The article body includes compact R and Python examples so the biological and scientific argument remains readable. The full repository expands those examples into a more rigorous computational cell-biology workflow, including exponential and logistic growth modeling, doubling-time estimation, viability-decay fitting, membrane diffusion and flux calculations, simplified cell-cycle transition models, treatment-response comparisons, cell-density normalization, imaging feature summaries, cell-condition scoring, SQL provenance structures, validation notes, reproducible data files, and full-stack scientific-computing examples across Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, and notebooks.
Systems cell biology and quantitative extension
Modern cell biology extends classical cell theory by treating the cell as a dynamic system rather than only a structural unit. Cellular behavior emerges from coupled networks of genes, proteins, lipids, metabolites, membranes, organelles, signaling pathways, mechanical forces, and environmental inputs. For that reason, cells are now studied not only by microscopy and biochemical assay but also through network analysis, high-content imaging, live-cell tracking, single-cell profiling, quantitative modeling, and computational inference.
This systems perspective is especially important in cancer biology, immunology, developmental biology, marine stress physiology, microbial ecology, pharmacology, and regenerative medicine. In these fields, cellular behavior often depends on interactions rather than on single components alone. A signaling pathway may change depending on nutrient state, membrane context, tissue architecture, or mechanical stress. A cancer cell may respond to treatment differently depending on microenvironment, immune interaction, and metabolic state. A phytoplankton cell may respond to warming differently depending on nutrients, light, salinity, pH, and viral pressure.
Quantitative cell biology makes it possible to estimate rates, fit models, compare perturbations, analyze high-dimensional datasets, and connect microscopy to molecular measurement. This strengthens rather than replaces classical cell theory. The cell remains the unit, but the unit is now understood as dynamic, networked, data-rich, and experimentally perturbable.
Systems cell biology also helps biology avoid a false choice between reduction and holism. Cells can be broken down into pathways, organelles, genes, and molecular interactions, but cellular behavior emerges from the organized relation among those parts. Cell theory remains foundational because it identifies the living system in which molecular mechanisms become coordinated biological function.
Limits and modern refinements
Cell theory remains foundational, but modern biology has refined its application. Some biological systems complicate simple cellular individuality: syncytia, multinucleated muscle fibers, fungal hyphae, plasmodesmata, gap junctions, bacterial biofilms, and colonial organisms all show that cells can be strongly interconnected while remaining meaningful biological units. Viral dependence on host cells also reminds us that cellular autonomy is central to life on Earth even where biologically consequential entities exist at the margins of cellular independence.
These complications do not overturn cell theory. They show that the cell is a basic unit, not the only relevant unit. Biological systems are multi-level. Cells build tissues, tissues build organs, organisms form populations, populations interact in ecosystems, and lineages evolve through time. The cell remains foundational because it is where many core processes of life are integrated, but modern biology must also study the relations among cells and the larger systems they compose.
Cell theory also requires caution in computational and biomedical contexts. Cell-line behavior in culture is not always equivalent to tissue behavior in vivo. Single-cell profiles require careful interpretation because dissociation, sampling, sequencing depth, batch effects, and analysis pipelines can influence results. Imaging features do not automatically explain mechanism. Growth models are simplified representations, not complete descriptions of cellular life. Modern cell theory is strongest when it joins cellular foundations with experimental humility.
This is especially important in biotechnology. A cultured cell line, engineered microbe, organoid, or synthetic circuit may be experimentally powerful, but it remains a model system with assumptions. Cell theory makes such systems intelligible; it does not make every cellular model automatically representative of the living world.
Why cell theory still matters
Cell theory still matters because it identifies one of biology’s most durable explanatory levels. Molecular biology explains mechanisms inside cells. Physiology explains coordinated function across cells. Developmental biology explains how cells generate form. Immunology explains how cells recognize and respond. Ecology explains how cellular organisms transform environments. Evolution explains how cellular lineages change. Biotechnology manipulates cells as platforms for production and discovery. Computational biology increasingly models cells as measurable systems.
The cell is therefore not only a textbook unit. It is a working unit of modern science. It is where genes are expressed, proteins are made, membranes regulate exchange, energy is transformed, pathogens interact, treatments act, development unfolds, stress is sensed, and biological continuity is reproduced. Even when biology studies whole organisms or ecosystems, cellular processes often mediate the mechanisms.
Cell theory also matters intellectually because it prevents life from becoming vague. It gives biology a concrete level at which living organization can be observed, tested, modeled, and compared. That is why it remains central from microscopy to genomics, from histology to single-cell sequencing, and from classical physiology to computational systems biology.
It also matters ethically and practically. Medicine, biotechnology, conservation, environmental monitoring, and public health all rely on cellular evidence. A society that understands cells better is better equipped to understand disease, ecological stress, biological repair, and the consequences of intervention in living systems.
Conclusion
Cell theory remains central to biology because it identifies the level at which living structure and function are fundamentally organized. Cells are the smallest systems that integrate boundary, metabolism, information, regulation, repair, response, and continuity in a biologically complete way. From nineteenth-century microscopy to modern molecular and systems biology, cell theory has provided a durable framework for understanding growth, heredity, physiology, disease, ecological function, and environmental response.
Its continued value lies in its extensibility. What began as a structural theory now includes membrane dynamics, organelle biology, signaling, cell-cycle control, apoptosis, differentiation, infection, immunity, ecological physiology, marine stress response, biotechnology, computational modeling, and networked cellular behavior. A modern scientist-facing account of cell theory should therefore treat the cell not only as the basic unit of life, but also as a measurable, modelable, and experimentally tractable unit of biological analysis across medicine, ecology, marine biology, and biotechnology.
Cell theory endures because it is both simple and expandable. Its central claim remains recognizable: life is cellular. But modern biology has shown how deep that claim becomes once cells are understood as dynamic, regulated, relational, data-rich systems embedded within organisms, environments, and evolutionary history.
Related articles
- Biology
- What Is Biology?
- Biology and the Scientific Understanding of Living Order
- The Rise of Modern Biological Thought
- Observation, Experiment, and the Methods of Biological Inquiry
- Classification, Taxonomy, and the Ordering of Life
- Life, Death, and the Problem of Biological Definition
- Water, Energy, and the Material Conditions of Life
- Biomolecules and the Chemical Basis of Life
- Cell Structure, Membranes, and Organelles
- Cell Signaling, Communication, and Biological Coordination
- Metabolism, Energy, and Biological Function
- Molecular Biology and the Flow of Genetic Information
- Systems Biology and the Logic of Biological Integration
Further reading
- Alberts, B. et al. (2002) Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK21054/
- Cooper, G.M. (2000) The Cell: A Molecular Approach. 2nd edn. Sunderland, MA: Sinauer Associates. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9839/
- Khan, Y.S. and Farhana, A. (2025) ‘Histology, cell’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK554382/
- Marine Biological Laboratory (n.d.) Theory of Cells as Basic Units of Life. Available at: https://www.mbl.edu/research/research-centers/eugene-bell-center/featured-projects/seeing-cells-exhibit/theory-cells-basic-units-life
- Nature Education (n.d.) What Is a Cell? Available at: https://www.nature.com/scitable/topicpage/what-is-a-cell-14023083/
- Nature Education (n.d.) Systems Biology of the Cell. Available at: https://www.nature.com/scitable/topicpage/systems-biology-of-the-cell-14458771/
- OpenStax (2018) ‘Studying cells’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/4-1-studying-cells
- OpenStax (2018) ‘Prokaryotic cells’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/4-2-prokaryotic-cells
- OpenStax (2018) ‘Eukaryotic cells’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/4-3-eukaryotic-cells
References
- Alberts, B. et al. (2002) Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK21054/
- Cooper, G.M. (2000) The Cell: A Molecular Approach. 2nd edn. Sunderland, MA: Sinauer Associates. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9839/
- Khan, Y.S. and Farhana, A. (2025) ‘Histology, cell’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK554382/
- Marine Biological Laboratory (n.d.) Theory of Cells as Basic Units of Life. Available at: https://www.mbl.edu/research/research-centers/eugene-bell-center/featured-projects/seeing-cells-exhibit/theory-cells-basic-units-life
- Nature Education (n.d.) Systems Biology of the Cell. Available at: https://www.nature.com/scitable/topicpage/systems-biology-of-the-cell-14458771/
- Nature Education (n.d.) What Is a Cell? Available at: https://www.nature.com/scitable/topicpage/what-is-a-cell-14023083/
- OpenStax (2018) ‘Eukaryotic cells’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/4-3-eukaryotic-cells
- OpenStax (2018) ‘Prokaryotic cells’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/4-2-prokaryotic-cells
- OpenStax (2018) ‘Studying cells’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/4-1-studying-cells