
Last Updated May 28, 2026
Life, death, and the problem of biological definition examine one of biology’s most difficult and enduring questions: what distinguishes living systems from nonliving matter, what counts as death in cells and organisms, and why biological definition becomes unstable at the margins of viruses, dormancy, sterility, symbiosis, synthetic systems, and evolutionary continuity. Biology can usually recognize familiar living beings with ease: bacteria, plants, fungi, animals, protists, and cells. Yet when the life sciences attempt to define life with precision, the problem becomes far more difficult. Many standard criteria—organization, metabolism, responsiveness, heredity, growth, homeostasis, reproduction, and Darwinian evolution—are real, but no single criterion resolves every borderline case.
This article develops Life, Death, and the Problem of Biological Definition as a foundational conceptual article within the Biology knowledge series. It treats life not as a mysterious essence or a simple checklist, but as a historically sustained organization of matter, energy, information, regulation, and evolutionary continuity. Living systems maintain organization while exchanging matter and energy with their surroundings. They repair, reproduce, develop, respond, metabolize, adapt, and participate in lineages. Death, likewise, is not merely the disappearance of matter. It is the breakdown of integrated biological process at one or more levels of organization.
Main Library
Publications
Article Map
Biology
Related Topic
Chemistry
Related Topic
Earth Science
Related Topic
Environmental Science
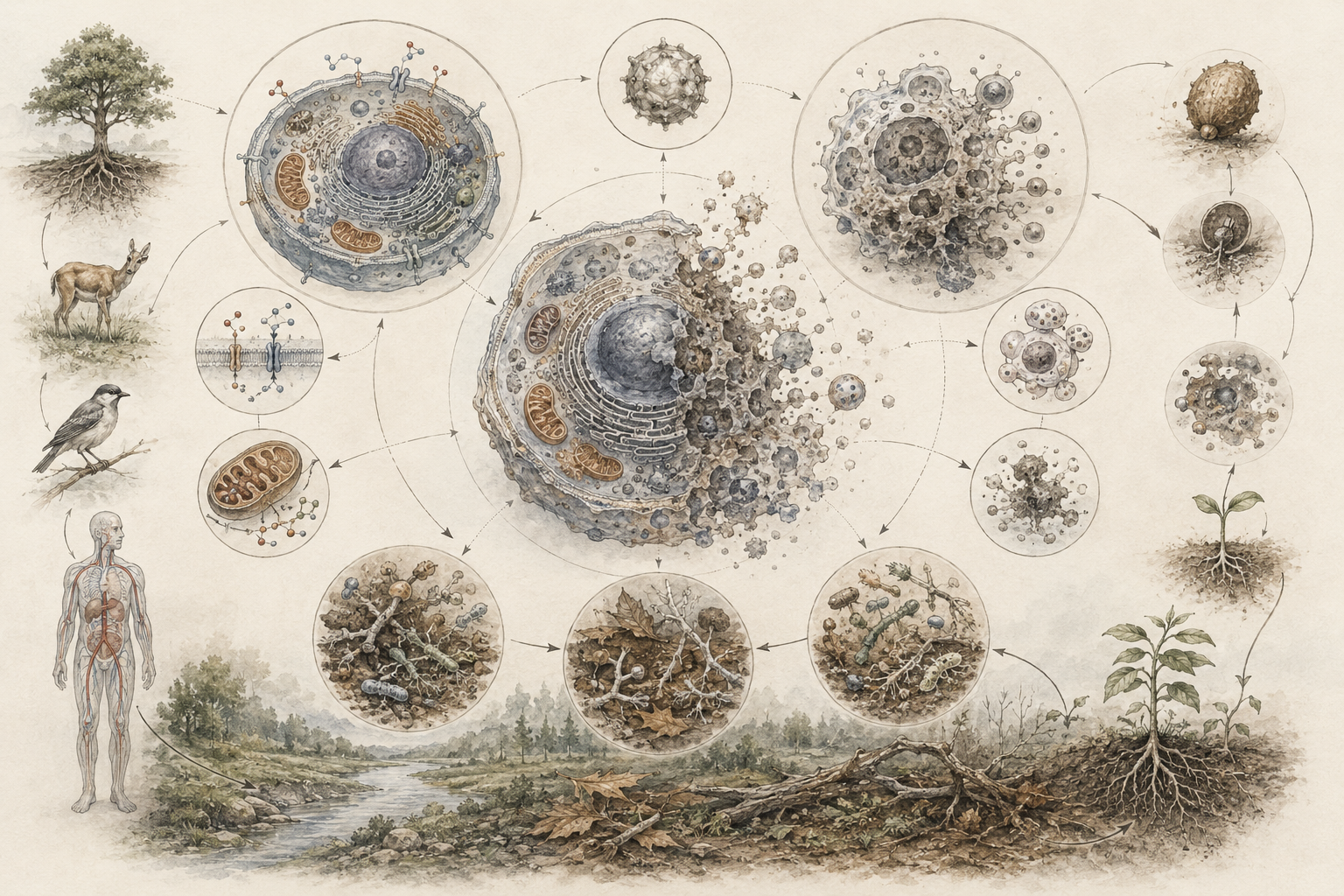
The article develops the definition problem across cells, organisms, lineages, viruses, dormant seeds and spores, bacterial persistence, programmed cell death, sterile organisms, symbioses, origin-of-life research, astrobiology, synthetic biology, medicine, ecology, marine systems, and computational biology. It emphasizes that life and death are not always best understood as simple binary categories. They often require attention to level, context, process, reversibility, autonomy, viability, and historical continuity.
The article also extends the problem into quantitative and computational biology through population growth, exponential viability decay, mortality hazard, dormancy-loss and reactivation models, host-virus dynamics, survival curves, borderline-case scoring, life-criteria matrices, R workflows, Python workflows, SQL provenance structures, and a linked full-stack GitHub repository containing Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, notebooks, data files, validation notes, and reproducibility documentation.
Why defining life is difficult
At first pass, life seems easy to identify. Biologists can usually distinguish a bacterium from a rock, a seedling from a crystal, or an animal from a machine. Yet when biology asks for a precise definition, the problem becomes much harder. Standard textbook traits such as metabolism, growth, responsiveness, heredity, and reproduction do not always appear together in every case, and they do not always separate living from nonliving systems without ambiguity.
A mule is alive though sterile. A seed can remain dormant for long periods with minimal detectable activity. A bacterial spore may persist through harsh conditions without obvious growth. A virus carries genetic information and evolves, yet does not maintain autonomous metabolism outside a host cell. Programmed cell death can be part of normal development rather than a simple biological failure. Some symbiotic systems complicate the very idea of what counts as an individual organism.
This difficulty arises because life is organized, historical, and variable. Living systems differ enormously in form, scale, metabolic intensity, reproductive mode, environmental dependence, and degree of autonomy. A human, a bacterium, a fungal spore, a phytoplankton cell, a coral colony, and a lichen are all biologically meaningful, yet they do not display “life” in the same way. Some definitions emphasize metabolism. Others emphasize heredity, bounded organization, autonomy, cellularity, or the capacity for Darwinian evolution. Each captures something real, but none captures everything.
The problem of biological definition is therefore not a semantic distraction. It exposes the difficulty of describing living organization in a way that is broad enough to cover life while still precise enough to exclude nonliving complexity. It also reveals that biology works with layered explanations: molecular, cellular, organismal, ecological, evolutionary, and planetary. Any definition of life that collapses these levels too quickly risks becoming either too narrow to be useful or too vague to be discriminating.
Classical hallmarks of life
Biology has often identified life through a recurring cluster of hallmarks: organized structure, metabolism, responsiveness to environmental conditions, growth or development, maintenance of internal order, heredity, reproduction, and participation in lineages that can evolve. These features remain useful because they capture much of what makes cells, plants, animals, fungi, and microbes biologically distinctive. They align with broader biological themes of organization, continuity, adaptation, regulation, and transformation.
Yet each criterion has limitations. Reproduction cannot be required in a simple individual sense because sterile organisms are still alive. Metabolism is central, but dormant forms can reduce activity dramatically. Responsiveness is widespread in life, yet certain nonliving systems also change state when environmental conditions change. Growth occurs in living systems, but crystals also grow in a looser physical sense. Homeostasis is important, but it appears differently in bacteria, plants, fungi, animals, and multicellular organisms with differentiated organs.
For that reason, the classical hallmarks function better as a family of indicators than as a perfectly decisive checklist. Their value lies not in reducing life to a single trait, but in identifying a cluster of processes that tend to occur together in living systems: organization, exchange, regulation, information, continuity, and evolutionary capacity.
This is one reason biology often proceeds pragmatically. Scientists may use a working definition that suits a particular context—virology, astrobiology, physiology, evolutionary biology, ecology, medicine, or synthetic biology—while recognizing that no single compact statement exhausts the phenomenon. The hallmarks of life remain useful, but their usefulness lies more in cumulative orientation than in absolute finality.
Life as process rather than thing
One influential response to the definition problem is to treat life less as a substance and more as a process. On this view, life is not a special material essence but a continuing organization of matter, energy, and information through regulation, exchange, repair, reproduction, and historically sustained continuity. A living organism remains itself while continuously turning over molecular components. What persists is not static matter, but organized dynamic pattern.
This process-centered perspective is scientifically useful because it aligns with contemporary biology. Organisms are not inert objects. They maintain internal order, interact with environments, and participate in lineages shaped by evolution. Cells regulate internal chemistry, organisms coordinate tissues and organs, and populations persist through reproduction and inheritance even as individuals die. In that respect, a process-based account of life fits better with modern physiology, cell biology, ecology, and systems biology than a rigid list of visible traits.
Thinking of life as process also helps explain why death is better understood as the breakdown of organized continuity than as the simple disappearance of matter. The atoms remain. What is lost is the integrated, self-maintaining pattern that made the system biologically active. That makes life and death less like the presence or absence of a substance and more like the maintenance or collapse of organized biological relation.
A process account does not eliminate the need for definitions. Instead, it shifts the question. Rather than asking whether life is one single property, it asks which processes must be present, at which level, and with what degree of autonomy, continuity, and evolutionary capacity. This makes the definition problem more precise.
Viruses and the problem of the borderline
Viruses are among the best-known challenges to simple definitions of life. They contain genetic material, evolve, adapt under selection, and participate profoundly in the history of life, yet they lack autonomous metabolism and cannot reproduce independently outside host cells. They therefore sit at the threshold between living and nonliving categories, or perhaps more precisely, at the threshold between autonomous and host-dependent biological organization.
Because of this, viruses occupy an ambiguous position. Some researchers describe them as nonliving infectious agents; others argue that the question is poorly framed because viruses are inseparable from the cellular life with which they coevolved. The broader point is not merely that viruses are difficult to classify. It is that biological definition often breaks down at the margins. Some entities sit at thresholds of autonomy, metabolism, heredity, and evolutionary agency, forcing biology to clarify which criteria matter most and why.
This ambiguity is not merely philosophical. Viral biology is central to ecology, marine systems, medicine, and biotechnology. Marine viral communities shape microbial mortality and nutrient cycling. Human viruses shape disease, immunity, and public health. Viral vectors are used in gene therapy and biotechnology. Phages shape bacterial populations and evolutionary dynamics. A conceptually difficult entity can still be experimentally central.
Viruses therefore reveal a key point: the boundary of life is not always best understood as a sharp line. It may be a zone of dependency, transition, and partial biological agency.
Dormancy, spores, seeds, and suspended life
The definition problem is also complicated by dormant forms. Seeds, spores, cysts, resting eggs, and other low-activity states may remain viable for long periods while showing sharply reduced metabolism. Biology does not treat them as dead because they retain organized potential and can resume active growth, metabolism, and reproduction under appropriate conditions. Dormancy therefore shows that life cannot be defined solely by visible activity at a single moment.
These cases reveal the temporal dimension of biological definition. To ask whether something is alive is not always to ask whether it is currently maximally active. It may also mean asking whether it belongs to an organized system capable of renewed biological process. Dormancy complicates rigid oppositions between life and death by introducing states of suspension, persistence, and reactivation.
Dormancy is also ecologically and medically important. Soil seed banks, microbial resting stages, plankton cysts, fungal spores, bacterial persistence, and latent infections all affect how living systems survive harsh conditions and re-emerge when circumstances improve. Life, in such cases, is less a constant visible activity than a retained capacity for organized revival.
This is especially important for environmental biology. Drought, freezing, heat, nutrient scarcity, oxygen limitation, and salinity can push organisms into states where activity is minimized but viability remains. The boundary between inactive life and biological death becomes a matter of reversibility, organization, and retained potential.
Sterility, symbiosis, and the problem of the individual
Sterility complicates definitions based too narrowly on reproduction. Individual organisms may be alive even when they cannot reproduce. Worker insects, mules, aged organisms, damaged organisms, and medically infertile organisms do not cease to be alive because they lack individual reproductive capacity. This suggests that reproduction should often be understood at the level of lineages, populations, or life cycles rather than as a necessary property of each individual.
Symbiosis complicates the problem further. Lichens, corals, microbiomes, endosymbiotic organelles, and host-associated microbial communities challenge the assumption that life is always organized into discrete, self-contained individuals. A coral depends on algal symbionts. A human body depends on microbial communities. Mitochondria and chloroplasts descend from formerly independent organisms. Biological individuality is therefore often relational, layered, and historically assembled.
The definition of life must be able to recognize these nested dependencies without dissolving every organism into an indistinct system. A cell, organism, colony, holobiont, population, and lineage may each be biologically relevant, but they are not identical units. The problem of definition is therefore also a problem of level.
A strong definition of life must therefore be flexible enough to recognize organismal integration and dependency at the same time. Living systems are often bounded, but not absolutely self-contained. They persist through exchange, relation, inheritance, and ecological embedding.
Death in biology
If life is difficult to define, death is equally complex. At one level, death marks the irreversible loss of integrated function. Yet biology also studies cell death, tissue death, programmed developmental death, and regulated mechanisms such as apoptosis. Death is therefore not always a single instant or a simple binary state. It can be a process of disintegration, a regulated biological mechanism, or a threshold defined differently at different levels of organization.
In multicellular organisms, cells can die while the organism lives. Tissues can fail while systemic life continues. Microbes may lose viability gradually across populations rather than all at once. Seeds may remain dormant but alive. Viruses may be inactivated without ever having been metabolically alive in the same sense as cells. These examples show that death must be interpreted relative to biological level and process.
This layered view is especially important in medicine, developmental biology, ecology, and conservation. Cell death is not merely the negation of life. In many cases it is part of how life regulates itself, shapes form, removes damaged cells, and preserves system-level integrity. Death, in biological terms, can therefore be both a terminal event and a functional process within living organization.
Death also matters ecologically. The death of one organism may become nutrient flow for another. Decomposition, predation, microbial metabolism, detrital food webs, and soil formation all show that biological death does not remove matter from living systems. It reorganizes matter through ecological process.
Cell death, programmed death, and biological function
Programmed cell death shows that death can be biologically functional. Apoptosis and other regulated cell-death pathways help shape development, eliminate damaged cells, maintain tissue organization, regulate immunity, and prevent uncontrolled growth. The death of a cell may support the life of the organism. This means death cannot be treated only as failure. It can be part of the organization of life.
This is crucial in developmental biology. The formation of tissues, organs, and body plans often depends on selective cell death. It is equally crucial in immunology, where infected, damaged, or abnormal cells may be eliminated to protect the system. In cancer biology, failure of appropriate cell death can contribute to pathological survival. In neurobiology, excessive or dysregulated cell death can contribute to degeneration.
The biological definition problem therefore becomes more subtle: life at one level can require death at another. A multicellular organism is not a simple aggregate of cells all maximizing survival independently. It is a regulated system in which cell proliferation, differentiation, repair, and death are coordinated.
This is one reason biology needs level-sensitive explanations. A cell may die while the tissue benefits. A tissue may be damaged while the organism survives. An organism may die while its genes persist through descendants. A population may lose individuals while the lineage continues. Life and death are biological realities, but they are not always located at one level.
Cells, organisms, populations, and different levels of life
Part of the definition problem arises because biology works across levels of organization. A cell is alive, but a multicellular organism is also alive as a higher-order integrated system. A lineage persists through generations, and a population evolves even though individual organisms die. Some definitions emphasize cellular autonomy, while others emphasize lineage-level evolutionary capacity. Still others stress organismal integration, homeostasis, or ecological embeddedness.
Biology therefore often works with nested rather than singular answers. Cells, organisms, populations, and lineages can all matter, but in different explanatory contexts. A cell biologist may treat the cell as the minimal living system. An evolutionary biologist may emphasize lineages and inheritance. An ecologist may focus on the persistence of populations and communities under environmental constraint. A medical scientist may focus on viability, reversible injury, and integrated organismal function.
This multi-level perspective also keeps biology from false simplicity. The same organism can be alive as an integrated whole while containing dying cells, symbiotic microbes, dormant states, inherited viral elements, and reproductive cells with different biological trajectories. Living systems are layered compositions, not single-level abstractions.
For that reason, definitions of life are often more useful when they clarify the level of analysis. A definition designed for astrobiology may differ from one used in clinical medicine, and both may differ from one used in evolutionary theory or synthetic biology. What matters is not only the definition itself, but the biological work it is being asked to perform.
Ecological, marine, and environmental relevance
The problem of life and death is especially relevant to ecology because ecological systems are structured by birth, growth, dormancy, mortality, decomposition, predation, parasitism, senescence, reproduction, and lineage persistence. Populations survive through reproduction, communities shift through colonization and extinction, and ecosystems recycle matter through death and decay. Ecological systems make clear that life is not an isolated property of individual organisms but part of larger cycles of persistence and transformation.
Marine biology adds another important dimension. In the oceans, life and death are often mediated through microbial loops, viral lysis, dormancy, nutrient release, predation, sinking organic matter, and extreme environmental gradients. Marine viruses shape plankton mortality and nutrient turnover. Resting stages allow organisms to survive low-resource or unfavorable conditions. Coral bleaching, hypoxia, warming, and acidification raise questions not only about organismal stress, but about the thresholds at which organized life breaks down.
Freshwater and soil systems show the same complexity. Dormant microbial pools, seed banks, fungal spores, biofilms, decomposition, and nutrient cycling all blur simple distinctions between active life, suspended life, dying matter, and recycled biological material. For ecologists, marine biologists, and environmental scientists, the question is not only “what is alive?” but “how does life persist, suspend itself, fail, transform, and re-enter ecological cycles under environmental constraint?”
This environmental perspective is especially important under climate change, biodiversity loss, pollution, and habitat transformation. Definitions of life and death become practical when they influence how scientists measure organismal stress, population decline, ecosystem collapse, restoration success, and biological recovery.
Medical, biomedical, and disease relevance
The problem of life and death is central to medicine and biomedicine. Clinical and biomedical practice depend on distinguishing viable from nonviable cells, reversible from irreversible injury, infection from colonization, dormancy from eradication, and organismal life from systemic failure. Cell death pathways such as apoptosis, necrosis, pyroptosis, ferroptosis, and other regulated responses matter directly for cancer biology, immunology, developmental biology, toxicology, and pathology.
Medical professionals encounter the biological definition problem in practical forms. What counts as irreversible loss of function? How should tissue viability be interpreted? How should latent infection be understood? How do viral agents occupy the boundary between autonomous and host-dependent biological activity? How should clinicians think about cells or organisms that are metabolically suppressed but potentially recoverable? These are not merely philosophical questions. They shape diagnosis, treatment, prognosis, public health, transplantation, oncology, infectious disease, and ethical decision-making.
Biomedicine therefore reinforces a key point: the boundary between life and death is often biologically real yet operationally complex. Living systems are dynamic, regulated, vulnerable, and layered. Any serious biological account of life must be able to speak to those realities.
This is why operational definitions matter. Clinical medicine, pathology, cell biology, and biotechnology may each require specific criteria for viability, death, persistence, or recovery. A single philosophical definition cannot replace those operational standards, but it can help clarify what they assume.
Life, origin, astrobiology, and the search beyond Earth
The difficulty of defining life becomes especially urgent in origin-of-life research and astrobiology. Scientists searching for life beyond Earth may encounter systems that do not resemble familiar terrestrial organisms. That means they need criteria disciplined enough to identify biologically meaningful organization, yet flexible enough not to miss unfamiliar forms. A definition that is too narrow may fail to recognize nonstandard life. A definition that is too broad may collapse the distinction between living and nonliving complexity.
This matters because the search for extraterrestrial life depends on criteria that are both rigorous and adaptable. If life on Earth already contains borderline cases such as viruses, dormancy, host-dependent replication, symbiosis, and low-activity states, then astrobiology cannot assume that extraterrestrial life will fit a simple classroom definition. In that sense, the problem of biological definition is not peripheral to planetary science. It is one of its conceptual foundations.
Origin-of-life research intensifies the problem further. Before life became fully established, there must have been transitions from chemistry to organized, self-maintaining, evolving systems. Any account of life must therefore be able to face not only the question of what living systems are now, but how organized biological continuity could have emerged from nonliving precursors.
This is where the definition problem becomes both empirical and philosophical. Scientists can study prebiotic chemistry, protocells, self-replicating systems, catalytic networks, lipid compartments, and metabolic-like cycles, but they must still ask when such systems cross from chemistry into biology. The answer may not be a single instant. It may be a transition zone in which biological features accumulate.
Biotechnology, synthetic biology, and operational definition
The definition problem also matters for biotechnology and synthetic biology. As biology gains the power to manipulate cells, genomes, pathways, viral vectors, cell-free systems, and engineered organisms, it increasingly confronts the question of what sort of organized entities it is designing, modifying, or controlling. Engineered microbes, gene-delivery vectors, synthetic circuits, minimal cells, organoids, and cell-free biochemical systems all push on the boundary between living organization and designed biological function.
Biotechnology depends on workable distinctions between viability, replication, persistence, host dependence, containment, and controlled activity. Synthetic biology raises especially sharp questions: if a system metabolizes, reproduces, evolves, or responds to the environment under engineered conditions, how should it be categorized? What distinguishes a programmable biological system from a merely biochemical one? Where does life begin in systems assembled through human intervention?
These questions do not make biotechnology philosophically vague. They make it conceptually serious. Modern biological engineering inherits the ancient question “what is life?” and must answer it in operational terms useful for design, regulation, biosafety, biosecurity, reproducibility, and ethical governance.
This is also why transparent biological definitions matter in applied settings. A regulator, engineer, physician, ecologist, or synthetic biologist may not need a final metaphysical theory of life, but they do need clear operational standards for what a system can do, how it persists, how it can fail, and how its risks should be governed.
Mathematical lens
The boundary between life and nonlife is partly conceptual, but several of the underlying processes can be studied quantitatively. Population growth, mortality, dormancy exit, host-virus interaction, viability loss, survival probability, and recovery potential are all measurable. That matters because the debate over life is not only philosophical. It also concerns processes such as replication, persistence, resilience, and breakdown that can be estimated from data.
A simple model for unconstrained population growth is:
Interpretation: Exponential growth describes increase in abundance when per-capita growth rate remains constant.
where \(N_0\) is the initial abundance, \(r\) is the per-capita growth rate, and \(t\) is time. A system that sustains growth and lineage continuity under appropriate conditions clearly differs from inert matter, even though growth alone is not a complete definition of life.
If viability declines in a stressed or dying population, a simple decay model is:
Interpretation: Viability decay models the loss of viable biological organization through time.
where \(L(t)\) is viable abundance at time \(t\), \(L_0\) is initial viable abundance, and \(k\) is a loss or mortality rate. This model does not define death philosophically, but it quantifies a process central to death: the loss of viable organized function through time.
A related survival model is:
Interpretation: Survival probability declines exponentially when the hazard rate is constant.
where \(S(t)\) is survival probability and \(h\) is a constant hazard rate. This is useful for viability assays, microbial persistence, toxicology, mortality studies, and ecological survival analysis.
A simple dormancy model is:
Interpretation: Dormant pools decline through mortality and reactivation, while the active pool increases through reactivation.
where \(D\) is the dormant pool, \(A\) is the activated pool, \(m\) is mortality during dormancy, and \(\alpha\) is the reactivation rate. This model is useful for microbial persistence, seed banks, plankton resting stages, fungal spores, and recovery dynamics after stress.
A standard within-host viral dynamics model is:
Interpretation: Host-virus dynamics formalize viral dependence on susceptible cells, infected-cell production, and clearance.
where \(T\) is susceptible target-cell abundance, \(I\) is infected-cell abundance, \(V\) is free-virus abundance, \(\beta\) is infection rate, \(\delta\) is infected-cell loss rate, \(p\) is virion production rate, and \(c\) is viral clearance rate. This system formalizes the dependency that makes viruses conceptually difficult: they reproduce and evolve, but only through host-cell machinery.
A transparent, heuristic life-criteria score can be written as:
Interpretation: A life-criteria score makes definitional assumptions explicit when comparing borderline cases.
where \(x_i\) are scaled criteria such as organization, metabolism, autonomy, heredity, responsiveness, and evolutionary capacity, while \(w_i\) are weights. Such a score is not a definition of life, but it can make assumptions explicit when comparing borderline cases.
Variables, units, and biological interpretation
Quantitative work on life, death, dormancy, and viability depends on variables that connect biological concepts to measurable processes. The table below summarizes several central quantities.
| Symbol or Term | Meaning | Typical Unit or Scale | Biological Interpretation |
|---|---|---|---|
| \(N(t)\) | Population or abundance at time \(t\) | cells, organisms, colonies, particles, or count proxy | Represents growth or persistence of a biological population |
| \(N_0\) | Initial abundance | same as \(N(t)\) | Starting condition for growth analysis |
| \(r\) | Per-capita growth rate | per unit time | Rate of increase under specified conditions |
| \(L(t)\) | Viable abundance at time \(t\) | viable cells, colony-forming units, seeds, spores, or survival count | Remaining viable biological units after stress, decay, or mortality |
| \(L_0\) | Initial viable abundance | same as \(L(t)\) | Starting viable pool before loss or stress exposure |
| \(k\) | Viability-loss or mortality rate | per unit time | Rate at which viable organization is lost |
| \(S(t)\) | Survival probability | probability from 0 to 1 | Probability that a biological unit remains viable or alive at time \(t\) |
| \(h\) | Hazard rate | per unit time | Instantaneous risk of death or failure under a model |
| \(D\) | Dormant pool | count or abundance | Biological units in a low-activity but potentially viable state |
| \(A\) | Activated pool | count or abundance | Dormant units that have re-entered active biological process |
| \(m\) | Dormancy mortality rate | per unit time | Rate of loss from the dormant pool due to death or irreversible damage |
| \(\alpha\) | Reactivation rate | per unit time | Rate at which dormant units become active |
| \(T, I, V\) | Target cells, infected cells, and free virus | counts or concentrations | State variables in host-virus dynamics |
| \(Q\) | Heuristic life-criteria score | dimensionless score | Transparent comparison scaffold for borderline cases |
The table shows why mathematical modeling can clarify the definition problem without solving it completely. A model can quantify growth, viability, dormancy, survival, or viral propagation, but interpretation still depends on level, context, and biological judgment.
Worked example: viability loss and dormancy retention
Suppose a stressed cell population begins with \(L_0=1.0\times10^6\) viable cells and declines to \(2.5\times10^5\) viable cells in 48 hours. Under the simple decay model:
Interpretation: Viability decay represents the decline of viable biological units over time.
we have:
Interpretation: The observed decline can be used to estimate the loss rate.
Dividing both sides by \(1.0\times10^6\):
Interpretation: Twenty-five percent of the initial viable population remains after 48 hours.
Solving for \(k\):
Interpretation: The estimated viability-loss rate is approximately 0.0289 per hour.
This parameter does not define death philosophically, but it gives a scientist a reproducible estimate of the rate at which viability is being lost under the experimental condition.
Dormancy can be formalized in a related way. Suppose a dormant pool begins with \(D_0=1.0\times10^6\), mortality during dormancy is \(m=0.02\), and reactivation rate is \(\alpha=0.05\). Then:
Interpretation: The dormant pool declines through both mortality and reactivation.
At \(t=20\):
Interpretation: After 20 time units, a substantial dormant pool remains viable or unactivated under the model assumptions.
The dormant pool declines, but it does not disappear immediately. This formalizes the idea that dormant life can persist through time as retained biological potential.
Computational modeling
Computational modeling helps make the life-definition problem more precise because many of the relevant processes are dynamic rather than static. A viability assay measures decline through time. Dormancy involves reversible and irreversible transitions. Host-virus dynamics depend on target cells, infected cells, and free viral particles. Survival analysis treats death or loss of viability as a probabilistic process. Borderline-case matrices make definitional assumptions visible rather than leaving them implicit.
The selected examples below focus on viability decay, life-criteria scoring, dormancy dynamics, host-virus dynamics, and survival curves. These examples are compact enough to appear in the article body, while the GitHub repository can extend them into more detailed simulations, sensitivity analyses, SQL provenance structures, validation tests, and multi-language scientific-computing workflows.
The purpose is not to define life by computation. The purpose is to use computation to clarify which processes are being measured, which assumptions are being made, and where conceptual judgment remains necessary.
R workflow: viability decay and life-criteria matrices
R is especially useful for viability analysis, statistical summaries, and transparent comparison tables. The following workflow estimates a viability-loss rate from live-cell counts and creates a heuristic matrix for comparing borderline cases.
# Viability Decay and Borderline Life-Criteria Matrix
#
# This workflow demonstrates two tasks related to biological definition:
#
# 1. Estimate viability-loss rate from live-cell counts.
# 2. Build a transparent heuristic comparison matrix for borderline cases.
#
# The life-criteria score is not a universal definition of life.
# It is a scaffold for making assumptions explicit.
library(tibble)
library(dplyr)
# ------------------------------------------------------------
# 1. Estimate viability-loss rate from live-cell counts
# ------------------------------------------------------------
viability_data <- tibble(
time_h = c(0, 12, 24, 36, 48),
live_cells = c(1.0e6, 7.4e5, 5.5e5, 3.8e5, 2.5e5)
)
# Linearized decay model:
# log(L) = log(L0) - k * t
decay_model <- lm(log(live_cells) ~ time_h, data = viability_data)
k_est <- -coef(decay_model)[["time_h"]]
L0_est <- exp(coef(decay_model)[["(Intercept)"]])
decay_summary <- tibble(
estimated_loss_rate_per_h = k_est,
estimated_initial_viable_count = L0_est,
half_life_h = log(2) / k_est,
r_squared_log_space = summary(decay_model)$r.squared
)
decay_fit <- viability_data %>%
mutate(
predicted_live_cells = exp(predict(decay_model)),
residual_log_scale = resid(decay_model)
)
# ------------------------------------------------------------
# 2. Heuristic life-criteria matrix
# ------------------------------------------------------------
cases <- tibble(
case = c("bacterium", "virus", "dormant_seed", "sterile_mule", "crystal"),
organization = c(0.95, 0.55, 0.80, 0.95, 0.35),
metabolism = c(0.90, 0.05, 0.20, 0.88, 0.00),
autonomy = c(0.88, 0.10, 0.45, 0.92, 0.00),
heredity = c(0.90, 0.82, 0.86, 0.80, 0.00),
responsiveness = c(0.85, 0.25, 0.40, 0.90, 0.05),
evolutionary_capacity = c(0.90, 0.88, 0.80, 0.20, 0.00)
)
weights <- c(
organization = 0.18,
metabolism = 0.18,
autonomy = 0.16,
heredity = 0.18,
responsiveness = 0.12,
evolutionary_capacity = 0.18
)
scored_cases <- cases %>%
mutate(
heuristic_life_score =
organization * weights[["organization"]] +
metabolism * weights[["metabolism"]] +
autonomy * weights[["autonomy"]] +
heredity * weights[["heredity"]] +
responsiveness * weights[["responsiveness"]] +
evolutionary_capacity * weights[["evolutionary_capacity"]]
) %>%
arrange(desc(heuristic_life_score))
print(round(decay_summary, 5))
print(round(decay_fit, 3))
print(round(scored_cases, 3))This R workflow is useful in cell-death assays, stress-response experiments, toxicology, microbial viability studies, environmental persistence work, and conceptual comparison of borderline cases. It turns raw viability counts into an interpretable parameter while making definitional assumptions explicit.
Python workflow: dormancy, host-virus dynamics, and survival curves
Python is especially useful for dynamic simulation, reproducible pipelines, and transparent biological modeling. The following workflow simulates dormancy loss and reactivation, simple host-virus dynamics, and a viability survival curve.
"""
Life, Death, Dormancy, and Host-Virus Dynamics
This workflow demonstrates three quantitative models related to
the problem of biological definition:
1. Dormancy loss and reactivation.
2. Host-virus dynamics.
3. Viability survival curves.
The examples are intentionally compact, but they can be extended
for seed banks, microbial persistence, plankton resting stages,
viral dynamics, toxicology, cell-death assays, and survival analysis.
"""
from __future__ import annotations
import numpy as np
import pandas as pd
def simulate_dormancy(
initial_dormant: float = 1.0e6,
mortality: float = 0.02,
reactivation: float = 0.05,
t_max: float = 20.0,
dt: float = 0.01,
) -> pd.DataFrame:
"""
Simulate dormancy loss and reactivation.
Dormant units leave the dormant pool through mortality and reactivation.
Reactivated units accumulate in the active pool.
"""
time = np.arange(0, t_max + dt, dt)
dormant = np.zeros_like(time)
active = np.zeros_like(time)
dead_or_lost = np.zeros_like(time)
dormant[0] = initial_dormant
for i in range(1, len(time)):
d_dormant = -(mortality + reactivation) * dormant[i - 1]
d_active = reactivation * dormant[i - 1]
d_lost = mortality * dormant[i - 1]
dormant[i] = max(dormant[i - 1] + d_dormant * dt, 0.0)
active[i] = active[i - 1] + d_active * dt
dead_or_lost[i] = dead_or_lost[i - 1] + d_lost * dt
return pd.DataFrame(
{
"time": time,
"dormant_pool": dormant,
"activated_pool": active,
"dead_or_lost_pool": dead_or_lost,
}
)
def simulate_host_virus(
initial_target_cells: float = 1.0e6,
initial_infected_cells: float = 0.0,
initial_free_virus: float = 1.0e3,
beta: float = 2e-8,
delta: float = 0.5,
production: float = 100.0,
clearance: float = 2.0,
t_max: float = 10.0,
dt: float = 0.01,
) -> pd.DataFrame:
"""
Simulate a simple target-cell, infected-cell, free-virus model.
"""
time = np.arange(0, t_max + dt, dt)
target_cells = np.zeros_like(time)
infected_cells = np.zeros_like(time)
free_virus = np.zeros_like(time)
target_cells[0] = initial_target_cells
infected_cells[0] = initial_infected_cells
free_virus[0] = initial_free_virus
for i in range(1, len(time)):
d_target = -beta * target_cells[i - 1] * free_virus[i - 1]
d_infected = (
beta * target_cells[i - 1] * free_virus[i - 1]
- delta * infected_cells[i - 1]
)
d_virus = production * infected_cells[i - 1] - clearance * free_virus[i - 1]
target_cells[i] = max(target_cells[i - 1] + d_target * dt, 0.0)
infected_cells[i] = max(infected_cells[i - 1] + d_infected * dt, 0.0)
free_virus[i] = max(free_virus[i - 1] + d_virus * dt, 0.0)
return pd.DataFrame(
{
"time": time,
"target_cells": target_cells,
"infected_cells": infected_cells,
"free_virus": free_virus,
}
)
def viability_survival_curve(
initial_viable_count: float = 1.0e6,
loss_rate: float = 0.0289,
t_max: float = 96.0,
n_points: int = 193,
) -> pd.DataFrame:
"""
Generate a simple exponential viability survival curve.
"""
time_h = np.linspace(0, t_max, n_points)
viable_count = initial_viable_count * np.exp(-loss_rate * time_h)
survival_probability = viable_count / initial_viable_count
return pd.DataFrame(
{
"time_h": time_h,
"viable_count": viable_count,
"survival_probability": survival_probability,
}
)
def main() -> None:
"""
Run compact models for dormancy, host-virus dynamics, and survival.
"""
dormancy_df = simulate_dormancy()
virus_df = simulate_host_virus()
survival_df = viability_survival_curve()
print("Dormancy simulation:")
print(dormancy_df.head(12).round(3).to_string(index=False))
print(dormancy_df.tail(12).round(3).to_string(index=False))
print("\nHost-virus simulation:")
print(virus_df.head(12).round(3).to_string(index=False))
print(virus_df.tail(12).round(3).to_string(index=False))
print("\nViability survival curve:")
print(survival_df.head(12).round(4).to_string(index=False))
print(survival_df.tail(12).round(4).to_string(index=False))
if __name__ == "__main__":
main()This Python workflow formalizes several of the article’s central claims. Dormant organisms or cells may persist in low-activity states without becoming biologically irrelevant. Viruses participate in measurable population processes that depend on host-cell availability, infected-cell loss, viral production, and clearance. Viability loss, death, and persistence can often be studied as time-dependent processes rather than single visual states.
GitHub repository
The article body includes compact R and Python examples so the biological and conceptual argument remains readable. The full repository expands those examples into a more rigorous computational life-definition workflow, including viability-decay fitting, mortality half-life estimation, dormancy-loss and reactivation models, host-virus dynamics, survival curves, life-criteria matrices, borderline-case scoring, SQL provenance structures, validation notes, reproducible data files, and full-stack scientific-computing examples across Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, and notebooks.
Limits, scaling, and modern biological definition
The problem of biological definition cannot be solved by a single model, metric, or checklist. A viability-decay model does not define death in every case. A host-virus model does not settle whether viruses are alive. A life-criteria score does not replace philosophical and biological analysis. These tools are useful because they make processes explicit, not because they eliminate conceptual difficulty.
Scaling is especially important. A cell may die while an organism lives. An organism may die while a lineage persists. A population may decline while dormant forms remain viable. A virus may be inert outside a host and active within a host-dependent cycle. A seed may appear inactive but retain organized potential. Different biological levels require different criteria.
Modern biology therefore benefits from plural definitions and operational clarity. In one context, life may be defined by cellular autonomy. In another, by Darwinian evolution. In another, by viable organization. In another, by biosignature evidence. In another, by clinical irreversibility. These are not always contradictions. They may be context-specific tools for different scientific tasks.
The strongest approach is therefore not to force every case into one rigid formula. It is to state the level of analysis, the criteria being used, the evidence available, and the reason those criteria matter for the scientific question at hand.
Why the definition problem matters
The problem of defining life matters because biology is not merely describing familiar organisms. It is also deciding how to interpret edge cases, how to classify unusual systems, and how to guide research in virology, ecology, medicine, synthetic biology, origin-of-life studies, and astrobiology. Whether viruses count as alive, how death is defined in complex organisms, and what criteria matter most for recognizing life have scientific, ethical, and institutional consequences.
The definition problem also matters philosophically because biology depends on concepts as well as measurements. A science can sequence genomes, observe cells, model ecosystems, and engineer biological systems while still needing to ask what kind of phenomenon “life” actually is. That is not a weakness of biology. It is a sign that life is a deep and difficult form of organized complexity.
Finally, the definition problem matters because it encourages intellectual humility. Biology can describe many hallmarks of life with confidence, yet the margins remain contested. That is not a failure of science. It is a reflection of the fact that life is richer than any one-line formula used to define it.
In that sense, the definition problem is not a temporary inconvenience that biology will eventually leave behind. It is one of the discipline’s permanent conceptual foundations.
Conclusion
Life, death, and the problem of biological definition show that one of biology’s most basic questions is also one of its most difficult. Living systems display recurring hallmarks such as organization, metabolism, responsiveness, heredity, regulation, and evolutionary continuity, yet no single criterion settles every case. Viruses, dormancy, sterility, regulated cell death, symbiosis, synthetic systems, and the transition from chemistry to biology all complicate rigid definitions.
The strongest biological approach is therefore often plural, process-oriented, and level-sensitive. Life is best understood not as a single checklist item but as a historically sustained organization of matter, energy, information, and evolutionary continuity. Death likewise is not always a single instant but often the breakdown of integrated biological process at one or more levels of organization.
Quantitative biology does not settle the conceptual problem by itself, but it helps formalize replication, persistence, dormancy, viability loss, mortality, survival, and host-dependent propagation in ways that make the debate scientifically sharper. To ask what life is, therefore, is to ask how biology understands organized complexity, continuity, vulnerability, and change. The question remains open not because biology knows nothing, but because life exceeds the simplicity of any one formula.
Related articles
- Biology
- What Is Biology?
- Biology and the Scientific Understanding of Living Order
- The Rise of Modern Biological Thought
- Observation, Experiment, and the Methods of Biological Inquiry
- Cell Theory and the Basic Unit of Life
- Water, Energy, and the Material Conditions of Life
- Biomolecules and the Chemical Basis of Life
- Cell Structure, Membranes, and Organelles
- Metabolism, Energy, and Biological Function
- Evolution and the History of Life
- Systems Biology and the Logic of Biological Integration
Further reading
- Alberts, B. et al. (2002) Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK21054/
- Cooper, G.M. (2000) The Cell: A Molecular Approach. 2nd edn. Sunderland, MA: Sinauer Associates. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9839/
- Koonin, E.V. and Starokadomskyy, P. (2016) ‘Are viruses alive? The replicator paradigm sheds decisive light on an old but misguided question’, Studies in History and Philosophy of Biological and Biomedical Sciences, 59, pp. 125–134. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5406846/
- NASA Astrobiology (n.d.) About Life Detection. Available at: https://astrobiology.nasa.gov/research/life-detection/about/
- NASA Astrobiology (n.d.) How can we tell if something is alive or not? Available at: https://astrobiology.nasa.gov/education/alp/alive-or-not/
- NASA Science (2023) Life on Other Planets: What is Life and What Does It Need? Available at: https://science.nasa.gov/universe/search-for-life/life-on-other-planets-what-is-life-and-what-does-it-need/
- National Research Council (2007) The Limits of Organic Life in Planetary Systems. Washington, DC: National Academies Press. Available at: https://nap.nationalacademies.org/catalog/11919/the-limits-of-organic-life-in-planetary-systems
- OpenStax (2018) ‘Homeostasis’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/1-3-homeostasis
- OpenStax (2018) ‘Themes and concepts of biology’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/1-introduction
References
- Alberts, B. et al. (2002) Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK21054/
- Cooper, G.M. (2000) The Cell: A Molecular Approach. 2nd edn. Sunderland, MA: Sinauer Associates. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9839/
- Koonin, E.V. and Starokadomskyy, P. (2016) ‘Are viruses alive? The replicator paradigm sheds decisive light on an old but misguided question’, Studies in History and Philosophy of Biological and Biomedical Sciences, 59, pp. 125–134. Available at: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5406846/
- NASA Astrobiology (n.d.) About Life Detection. Available at: https://astrobiology.nasa.gov/research/life-detection/about/
- NASA Astrobiology (n.d.) How can we tell if something is alive or not? Available at: https://astrobiology.nasa.gov/education/alp/alive-or-not/
- NASA Science (2023) Life on Other Planets: What is Life and What Does It Need? Available at: https://science.nasa.gov/universe/search-for-life/life-on-other-planets-what-is-life-and-what-does-it-need/
- National Research Council (2007) The Limits of Organic Life in Planetary Systems. Washington, DC: National Academies Press. Available at: https://nap.nationalacademies.org/catalog/11919/the-limits-of-organic-life-in-planetary-systems
- OpenStax (2018) ‘Homeostasis’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/1-3-homeostasis
- OpenStax (2018) ‘Themes and concepts of biology’, in Biology 2e. Available at: https://openstax.org/books/biology-2e/pages/1-introduction