
Last Updated May 28, 2026
Metabolism, energy, and biological function examine how living systems acquire, transform, store, allocate, and use energy in order to maintain organization, grow, reproduce, repair damage, respond to environments, and persist through time. Metabolism is one of the most fundamental principles in biology because no living system remains alive without continuous energetic and material throughput. A genome may encode possibility, a membrane may define boundary, and a tissue may provide structure, but life remains active only because matter and energy move through organized biochemical networks. Metabolism is therefore not merely a collection of pathways. It is the living system’s economy of transformation.
This article develops Metabolism, Energy, and Biological Function as a foundational article within the Biology knowledge series. It treats metabolic life not only as cellular chemistry, but as the framework through which energy flow becomes biological function across molecules, cells, organisms, populations, ecosystems, and planetary cycles. Metabolism links ATP and redox chemistry to physiology, photosynthesis to food webs, microbial respiration to biogeochemical cycling, plant growth to carbon storage, oxygen limitation to disease and ecosystem stress, and computational modeling to reproducible biological inference.
Main Library
Publications
Article Map
Biology
Related Topic
Chemistry
Related Topic
Earth Science
Related Topic
Environmental Science
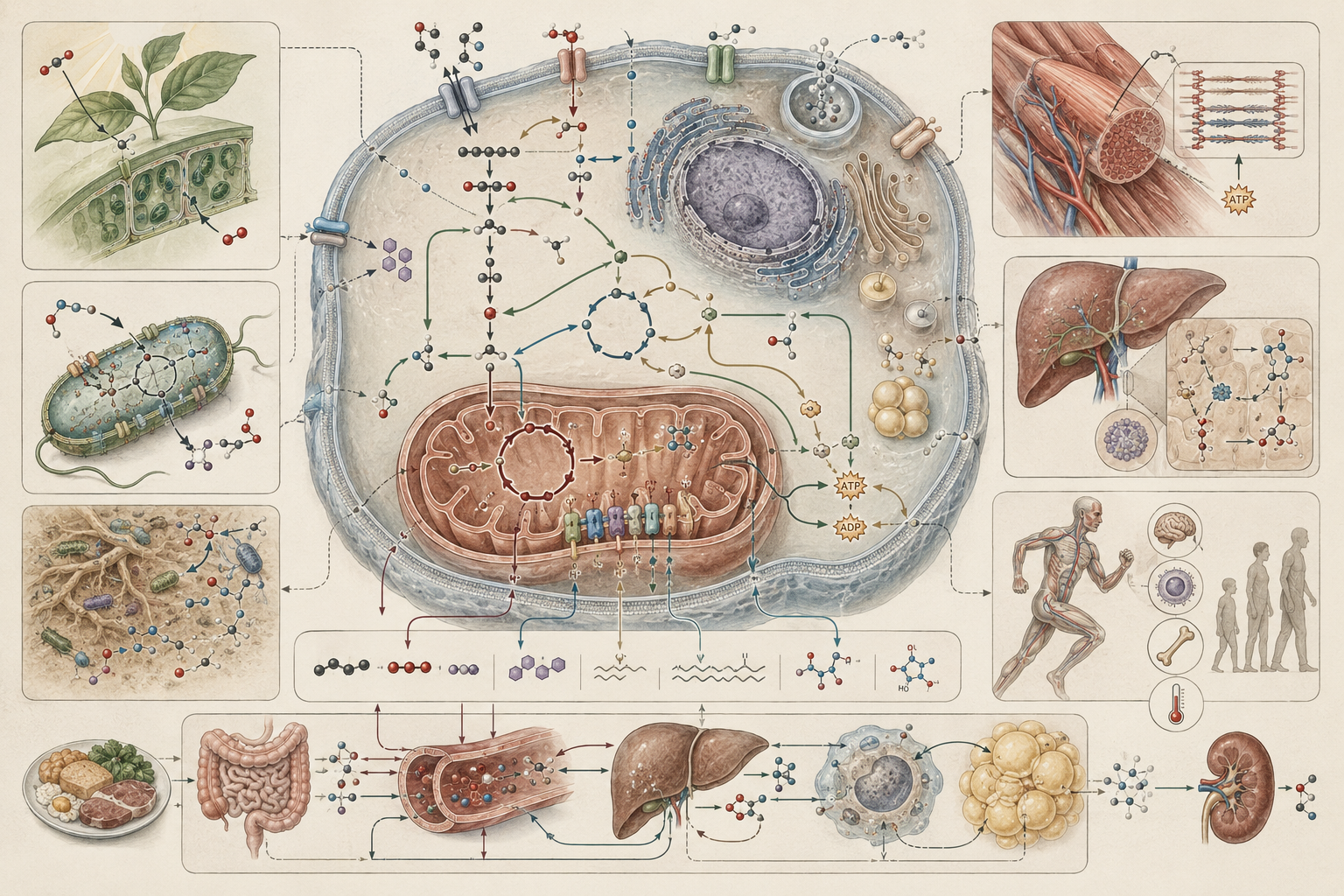
The article develops metabolism, energy, and biological function across catabolism, anabolism, ATP, redox carriers, cellular respiration, oxidative phosphorylation, fermentation, photosynthesis, primary production, metabolic regulation, growth, biomass yield, substrate uptake, oxygen consumption, energy allocation, thermodynamic constraint, microbial metabolism, plant metabolism, disease metabolism, biotechnology, metabolic engineering, ecological metabolism, and systems biology.
The article is written for biologists, biochemists, microbiologists, plant scientists, physiologists, ecologists, medical and environmental-health readers, biotechnology practitioners, computational biology readers, systems biologists, and sustainability-oriented researchers who need a rigorous account of how energy and matter become organized biological function across scales.
The article also extends metabolism into quantitative and computational biology through exponential and logistic growth models, Monod-style substrate limitation, biomass yield, maintenance allocation, oxygen consumption, ATP accounting, redox balance, simplified pathway flux, toy flux-balance analysis, energy-budget modeling, respirometry summaries, metabolic condition scoring, R workflows, Python workflows, SQL provenance structures, and a linked full-stack GitHub repository containing Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, notebooks, data files, validation notes, and reproducibility documentation.
What Metabolism Is
Metabolism is the totality of chemical reactions through which living systems maintain themselves, acquire usable energy, build and repair structures, eliminate wastes, regulate internal conditions, and perform biological work. In the broadest sense, metabolism is not one pathway but a coordinated network of transformations that links matter, energy, information, and function. Every cell depends on metabolism because no living organization can persist without continuous chemical work.
This is one reason metabolism sits near the center of biology. A cell membrane, genome, organelle, or tissue architecture may define aspects of living structure, but none remains biologically active without metabolic throughput. Molecules must be synthesized and degraded, gradients must be maintained, nutrients must be imported or generated, wastes must be processed, and energy must be stored in forms that can be mobilized for cellular work. A living system is therefore not just an organized structure. It is an organized flux.
Metabolism also reveals that life is inseparable from environment. Organisms do not create energy from nothing. They obtain matter and energy from sunlight, food, chemical gradients, prey, hosts, detritus, symbionts, or other environmental sources. Metabolic life is therefore always relational. It exists through exchange with surroundings, and that exchange connects cells to organisms, organisms to ecosystems, and ecosystems to planetary cycles.
This relational character is especially important for modern biology. Metabolism explains why cells must be studied in context: oxygen availability, nutrient status, temperature, pH, hydration, redox state, substrate limitation, waste accumulation, and ecological interaction all influence what metabolism can do. A pathway diagram is useful, but metabolism in a living system is never just a diagram. It is pathway behavior under constraint.
Energy, Life, and the Maintenance of Order
Living systems must constantly oppose local tendencies toward disorder by maintaining organized structure through regulated input and expenditure of energy. This does not violate thermodynamic law; it expresses it biologically. Cells and organisms remain ordered not by escaping physical constraint but by channeling energy through controlled pathways that permit repair, growth, transport, signaling, and internal regulation. Metabolism is therefore one of the main ways life maintains order under conditions of flux.
This is why energy is not merely one topic among others in biology. It is integral to nearly every biological function. Muscle contraction, membrane transport, protein synthesis, detoxification, neural signaling, immunity, development, reproduction, and environmental response all depend on energy transformation. Even apparently static structures are metabolically costly to maintain. A living system that stops processing energy does not remain alive for long.
From a broader perspective, metabolism links scales of life. The energetic logic that governs ATP production in a single cell connects to the physiology of an organism, the productivity of an ecosystem, and the circulation of carbon and nutrients across the biosphere. Biology therefore understands energy not only as a cellular issue, but as one of the great connective principles of life.
Energy also clarifies vulnerability. When energy supply fails, living order can deteriorate quickly. Ion gradients collapse, transport fails, repair slows, signaling becomes distorted, and cell death pathways may activate. Conversely, when energy is abundant but poorly regulated, organisms can suffer oxidative stress, metabolic disease, inflammatory dysregulation, or ecological imbalance. Biological function depends not only on access to energy, but on regulated use of energy.
Catabolism, Anabolism, and Metabolic Organization
Metabolism is commonly divided into two interacting domains: catabolism and anabolism. Catabolic processes break down molecules and, in doing so, release energy, harvest electrons, and generate intermediates that can be used for subsequent work. Anabolic processes build cellular components, macromolecules, tissues, storage compounds, and reproductive material, and these constructive activities require energetic input. The distinction is conceptually useful, but in living systems catabolism and anabolism are tightly coupled rather than isolated.
Catabolism provides more than raw energy. It generates reducing power, precursor metabolites, carbon skeletons, and molecular substrates that feed into biosynthesis. Anabolism, in turn, converts metabolic inputs into proteins, lipids, nucleic acids, membranes, structural carbohydrates, storage products, secondary metabolites, pigments, and countless other components of living organization. A cell is alive partly because these reactions are coordinated rather than merely occurring side by side.
This coupling is one of metabolism’s central features. Living systems do not simply burn fuel. They transform matter and energy through ordered networks in which breakdown and synthesis, release and capture, degradation and construction are mutually dependent. Biological function emerges not from one pathway but from the structured integration of many pathways.
Metabolism is therefore a systems problem from the beginning. It requires attention to flux, regulation, allocation, bottlenecks, feedback, and context. A metabolic pathway may appear linear on a page, but inside living systems it is embedded in networks of allosteric regulation, compartmentalization, enzyme abundance, substrate availability, redox balance, energy charge, gene expression, and environmental constraint.
ATP, Redox, and the Cellular Economy of Energy
Adenosine triphosphate, or ATP, is often described as the cell’s primary energy currency because it stores free energy in a chemically accessible form that can be coupled to work. ATP hydrolysis helps drive biosynthesis, transport, motility, signaling, protein folding, and regulation, while ATP regeneration requires energy harvested from catabolism, photosynthesis, chemolithotrophy, or other metabolic processes. ATP is not “energy” itself in the abstract. It is a molecular mechanism for coupling favorable and unfavorable processes within living systems.
ATP is only part of the story. Metabolism also depends on redox chemistry. Electrons are transferred through pathways involving carriers such as NADH, NADPH, and FADH2, and these electron transfers allow cells to couple oxidation and reduction to energy conservation, biosynthesis, detoxification, antioxidant defense, and signaling. In this sense, metabolism is an economy of both phosphate-bond transfer and electron flow.
The importance of this energetic economy is difficult to overstate. Cells survive because they can couple exergonic and endergonic reactions with sufficient precision to maintain internal order, build structure, respond to change, and preserve viability. Energy metabolism is therefore not simply about fuel. It is about the regulated conversion of chemical potential into biological function.
This also explains why metabolic failure can be so damaging. If ATP regeneration, redox balance, or substrate flow is disrupted, the consequences propagate through membranes, organelles, transcription, signaling, repair, immunity, and survival. Mitochondrial dysfunction, hypoxia, ischemia, toxin exposure, starvation, sepsis, metabolic syndrome, and environmental stress all demonstrate that the cell’s energy economy is a whole-system issue.
Cellular Respiration and the Harvesting of Energy
Cellular respiration is one of the major metabolic frameworks through which organisms harvest usable energy from organic molecules. In aerobic organisms, glucose and other carbon substrates are broken down through glycolysis, the citric acid cycle, and oxidative phosphorylation, yielding ATP and reduced electron carriers while also supplying intermediates for biosynthesis. Respiration is therefore both an energy-harvesting system and a material-routing system.
Respiration matters because it shows how efficiently biology organizes stepwise oxidation. Rather than releasing all energy at once, cells channel energy through controlled reactions that allow conservation in ATP and electrochemical gradients. Oxidative phosphorylation is especially important because it couples electron transfer to proton-motive force and ATP synthesis, thereby converting redox potential into broadly usable cellular energy.
Yet respiration is not one universal template. Different organisms rely on different substrates, electron donors, electron acceptors, and environmental conditions. Anaerobic pathways, fermentation, nitrate respiration, sulfate reduction, methanogenesis, and chemolithotrophic metabolism all show that metabolism is diverse, not monolithic. This diversity is biologically decisive because energetic strategy often shapes ecological role, environmental tolerance, evolutionary possibility, and biogeochemical consequence.
Cellular respiration therefore belongs to both cell biology and ecology. It is a molecular pathway, an organismal capacity, and an ecosystem process. Oxygen consumption in a mitochondrion, respiration in a microbial mat, decomposition in soil, and oxygen depletion in a lake are not the same phenomenon at the same scale, but they are connected by the metabolic transformation of matter and energy.
Photosynthesis, Primary Production, and the Entry of Energy into Living Systems
If respiration exemplifies energy harvesting from organic molecules, photosynthesis exemplifies the capture of external energy and its entry into living systems. Photosynthetic organisms transform light energy into chemical energy, producing ATP, reducing power, and carbon-rich compounds that sustain not only their own growth but much of the biosphere’s food-web structure. In that sense, metabolism is not only about expenditure and maintenance. It is also about the primary entry of energy into ecological systems.
Plant metabolism, algal metabolism, and microbial photosynthesis are central to sustainability-adjacent biology. Primary production supports terrestrial food systems, marine productivity, freshwater ecosystems, carbon storage, oxygen dynamics, and global biogeochemical cycling. Photosynthetic metabolism also links cell biology to larger-scale ecological and climate processes, since changes in productivity alter carbon sequestration, food availability, atmospheric exchange, and ecosystem resilience.
This is one reason metabolism must be understood across scales. ATP and chloroplast chemistry matter, but so do crop yields, plankton blooms, forest productivity, wetland carbon storage, and land-atmosphere exchange. Metabolism is biologically local and ecologically planetary at the same time.
Photosynthesis also reveals metabolism as constraint-sensitive. Light, water, nutrients, temperature, CO2, stomatal regulation, salinity, drought, and photodamage all shape photosynthetic performance. A plant or phytoplankton cell does not simply “perform photosynthesis” in isolation. It performs photosynthesis under material and environmental conditions that determine rate, allocation, and resilience.
Fermentation, Anaerobic Metabolism, and Metabolic Diversity
Fermentation and anaerobic metabolism show that life has many ways to maintain energy flow under oxygen limitation or absence. Fermentation allows cells to regenerate redox balance and produce some ATP without using oxygen as a terminal electron acceptor. Anaerobic respiration uses alternative electron acceptors such as nitrate, sulfate, carbon dioxide, iron, or other compounds depending on organism and environment. These pathways are not primitive curiosities. They are central to microbial ecology, food systems, biotechnology, sediments, wetlands, animal guts, wastewater treatment, and global biogeochemical cycles.
This metabolic diversity matters because environmental context determines which energetic strategies are viable. Oxygen-rich tissues, oxygen-poor sediments, stratified lakes, flooded soils, animal digestive tracts, biofilms, hydrothermal systems, and industrial bioreactors all create different metabolic possibilities. Microbial life in particular is defined by extraordinary diversity in electron donors, electron acceptors, carbon sources, and energy-conservation strategies.
Fermentation is also practically important. Bread, yogurt, cheese, beer, wine, vinegar, fermented vegetables, biofuels, industrial enzymes, organic acids, and many bioprocesses depend on controlled metabolic transformations. Biotechnology therefore uses metabolism not only as a natural phenomenon but as a production platform.
Anaerobic metabolism also connects to disease and environmental risk. Tissue hypoxia alters cellular metabolism. Tumor microenvironments can favor altered metabolic states. Wetlands and ruminant digestion contribute to methane production. Eutrophication and oxygen depletion can shift aquatic systems toward anaerobic pathways. Metabolic diversity is therefore scientifically powerful because it connects chemistry, ecology, medicine, and applied systems.
Metabolism, Physiology, and Biological Function
Metabolism underlies physiology because biological function depends on energetic supply, molecular turnover, and regulated exchange. A tissue cannot function without substrates, oxygen or alternative electron acceptors, ion gradients, and ATP turnover. Endocrine signaling, thermoregulation, muscle work, neural activity, renal function, immune response, digestion, reproduction, and development all require specific metabolic support.
Metabolism also helps explain why physiological failure can propagate quickly. If energy production is interrupted, transport fails, gradients collapse, signaling becomes unreliable, repair slows, and structural maintenance deteriorates. This is true not only in severe pathology but also in environmental stress, oxygen limitation, toxin exposure, nutrient deprivation, thermal stress, infection, or organismal overexertion.
From the standpoint of biological function, metabolism is therefore not background chemistry. It is one of the main ways biological systems turn material throughput into organized performance. A functioning organism is not simply a body with parts. It is an energy-transforming, matter-exchanging, regulated system that remains viable through continuous metabolic work.
This is especially clear in high-demand systems. The brain depends on continuous energy supply. Muscles shift metabolic strategy with activity. Immune cells change metabolism during activation. Plants allocate carbon among growth, storage, defense, roots, shoots, and reproduction. Microbes alter metabolism depending on substrate, oxygen, and stress. Physiology is metabolism organized into function.
Ecology, Biogeochemical Cycles, and Sustainability-Adjacent Biology
Metabolism is central to ecology because ecological systems are structured by the transformation and circulation of energy and matter through living beings. Food webs are metabolically linked. Nutrients are biologically assimilated, released, transformed, and recycled. Productivity, decomposition, respiration, trophic transfer, symbiosis, predation, and detrital processing are all metabolic processes expressed at population, community, and ecosystem scales.
This is where metabolism becomes especially relevant to sustainability-adjacent biology. Carbon cycling, nitrogen transformation, phosphorus limitation, decomposition, soil fertility, oxygen dynamics, methane production, microbial remediation, and plant productivity all depend on metabolic processes. Sustainability is therefore not only about managing resources abstractly. It is about understanding how living systems transform the material basis of survival under conditions of environmental limit, disturbance, and long-term change.
Ecology is central here because it reveals metabolism as relation rather than isolated chemistry. An organism’s metabolic strategy shapes its niche, resilience, and interactions, while ecosystem conditions constrain what kinds of metabolism are viable. Biology, in this sense, becomes a science of energetic interdependence.
This perspective also clarifies ecological collapse and recovery. A degraded ecosystem is not only a loss of species. It is often a disruption of metabolic processes: primary production, decomposition, nutrient retention, microbial respiration, oxygen balance, carbon storage, or trophic transfer. Restoration ecology therefore depends partly on restoring functional metabolic processes across living systems.
Marine, Freshwater, Soil, Plant, and Fungal Relevance
Marine biology makes metabolic interdependence especially visible. Marine primary production, microbial loops, viral lysis, oxygen minima, nutrient limitation, and carbon export all depend on the metabolic activities of cells and organisms distributed through water columns and food webs. Metabolism influences phytoplankton growth, coral symbiosis, fisheries productivity, marine microbial metabolism, and the response of marine organisms to warming, acidification, and hypoxia.
Freshwater biology presents parallel dynamics in rivers, lakes, wetlands, and aquifers, where metabolism shapes nutrient cycling, oxygen depletion, algal blooms, microbial decomposition, and ecosystem response to pollution or eutrophication. Aquatic metabolism is often visible through oxygen dynamics because photosynthesis, respiration, decomposition, and mixing shape whether systems remain productive or become stressed.
Soil biology likewise depends heavily on metabolism, since microbes, fungi, roots, and detrital communities drive decomposition, mineralization, nutrient release, carbon stabilization, and the maintenance or loss of fertility. Soil metabolism is not a metaphor. It is a distributed biochemical reality that links microbial activity, fungal networks, plant roots, organic matter, water, oxygen, and minerals.
Plant science and agroecology are deeply metabolic as well. Photosynthesis, respiration, nutrient uptake, growth allocation, water stress, nitrogen assimilation, carbon partitioning, seed filling, and root-microbe interaction all involve metabolic tradeoffs that shape food systems and land use. Forestry and restoration ecology depend on the same logic because recovery, resilience, and carbon storage are inseparable from how living systems process energy and matter.
Fungal biology adds a further dimension. Fungi metabolize complex organic matter, form mutualisms with plants, redistribute nutrients through mycelial networks, and drive decomposition in forests, soils, and agricultural systems. Their metabolic capacities are central to carbon cycling, soil formation, plant health, and biotechnological applications.
Medical, Biomedical, and Disease Ecology Relevance
Metabolism is foundational to medicine and biomedicine because health and disease depend on energy transformation, substrate use, redox balance, and metabolic regulation. Cancer biology, endocrinology, immunology, neurology, cardiovascular physiology, exercise physiology, mitochondrial disease, infectious disease, and critical care all require metabolic interpretation. Tissue viability depends on oxygen delivery, ATP production, redox balance, substrate flow, and waste handling.
Cells under stress may reprogram metabolism, pathogens may exploit host metabolic systems, and immune cells alter energetic pathways as they activate, differentiate, and respond. Disease ecology also depends on metabolic context because pathogens, hosts, vectors, microbiomes, and environments all operate under energetic and nutritional constraints. This is one reason metabolism can connect biomedical thinking to ecology rather than separating them.
Medical professionals therefore do not encounter metabolism only in textbook biochemistry. They encounter it in tissue viability, fatigue, inflammation, hypoxia, insulin signaling, mitochondrial dysfunction, infection, wound healing, organ failure, exercise tolerance, and recovery. Metabolism is not peripheral to function. It is one of the main ways biological possibility becomes clinically real.
This also explains why metabolic biomarkers are so important. Glucose, lactate, oxygen saturation, pH, ketones, lipids, creatinine, inflammatory metabolites, endocrine signals, mitochondrial markers, and metabolomic profiles can indicate the energetic and biochemical state of tissues or organisms. Metabolism provides a language for reading biological stress and adaptation.
Biotechnology, Bioinformatics, and Computational Relevance
Biotechnology extends metabolism into applied systems of measurement, control, and design. Fermentation, metabolic engineering, microbial production, environmental biotechnology, industrial bioprocessing, synthetic biology, wastewater treatment, biosensor development, and bioremediation all depend on understanding how cells allocate resources, channel substrates, and convert chemical energy into biomass, products, or ecological effects. In biotechnology, metabolism becomes not only a subject of explanation but also a target of intervention.
This is why computational approaches have become so important. Metabolic pathways can be modeled as networks, fluxes can be estimated, yields can be compared across conditions, growth curves can be fitted, oxygen consumption can be summarized, resource allocation can be simulated, and pathway bottlenecks can be inferred from data. R and Python support these workflows through statistical modeling, simulation, automation, and reproducible analysis.
Bioinformatics and systems biology extend this further. Genome-scale metabolic reconstructions, flux-balance analysis, constraint-based modeling, isotope tracing, transcriptomics, proteomics, metabolomics, and kinetic modeling all help connect molecular components to pathway behavior. The question is not merely how metabolism works in principle, but how metabolic behavior can be quantified and compared under real experimental, ecological, biomedical, and applied conditions.
Environmental biotechnology makes the sustainability link especially strong. Pollution breakdown, wastewater treatment, fermentation, bioremediation, soil recovery, methane mitigation, nutrient recovery, and biosensor development all depend on metabolic capacities distributed across microbial and multicellular systems. Metabolism is therefore one of the clearest bridges from basic biology to applied sustainability work.
Mathematical Lens
Modern metabolism research is not only descriptive and mechanistic. It is also quantitative. Growth rates, substrate uptake, product yield, ATP turnover, oxygen consumption, nutrient limitation, maintenance costs, redox balance, and pathway flux can all be studied mathematically and computationally. This does not replace biological interpretation. It makes interpretation sharper, more explicit, and more reproducible.
For a population growing under unconstrained early conditions, one simple approximation is:
Interpretation: Exponential growth describes abundance increase when per-capita growth rate remains approximately constant.
where \(N_0\) is the initial abundance, \(r\) is the per-capita growth rate, and \(t\) is time. Under resource limitation, logistic growth is often more realistic:
Interpretation: Logistic growth represents abundance increase under carrying-capacity or resource limitation.
where \(K\) is carrying capacity. These models are useful because metabolism and growth are tightly coupled: cells and organisms cannot increase biomass without energetic and material throughput.
A Monod-style growth relation can be written as:
Interpretation: Substrate-limited growth increases with substrate availability but approaches a maximum rate.
where \(\mu(S)\) is substrate-dependent growth rate, \(\mu_{\max}\) is maximal growth rate, \(S\) is substrate concentration, and \(K_s\) is the half-saturation constant. This is useful because microbial growth, algal production, fermentation, and nutrient limitation often depend on substrate availability.
A simple biomass yield relation can be written as:
Interpretation: Biomass yield measures how efficiently consumed substrate is converted into biomass.
where \(Y_{X/S}\) is biomass yield on substrate, \(\Delta X\) is change in biomass, and \(\Delta S\) is substrate consumed. This is useful because metabolism is often assessed not only by how fast organisms grow, but by how efficiently they convert resources into structure and function.
A simple allocation relation can be written as:
Interpretation: Input substrate may be allocated to growth, maintenance, product formation, or loss.
This is useful because metabolism often involves tradeoffs. Under stress, a larger fraction of substrate may be allocated to maintenance, repair, detoxification, osmotic regulation, or survival rather than growth.
Transport across membranes or gradients can be approximated with Fick’s first law:
Interpretation: Diffusive flux follows the concentration gradient and is scaled by the diffusion coefficient.
where \(J\) is flux, \(D\) is diffusion coefficient, and \(\frac{dC}{dx}\) is the concentration gradient. This becomes especially relevant in nutrient uptake, oxygen delivery, microbial mats, soil gradients, aquatic systems, and transport-limited assay design.
A compact pathway-flux approximation may be written as:
Interpretation: Pathway throughput can be constrained by the slowest or most limiting step.
where \(J\) is simplified pathway throughput and \(v_i\) are step-specific capacities. This is an oversimplified but useful teaching model because pathway throughput can be constrained by bottleneck steps rather than by average pathway capacity.
In simplified constraint-based metabolic modeling, steady-state mass balance is often written as:
Interpretation: At steady state, internal metabolites are balanced by production and consumption fluxes.
where \(S\) is a stoichiometric matrix and \(v\) is a vector of reaction fluxes. This is useful because it expresses the idea that, at steady state, internal metabolites are balanced by production and consumption fluxes.
Variables, Units, and Metabolic Interpretation
Quantitative metabolism depends on variables that connect abundance, growth, substrate use, energy allocation, flux, yield, and biological interpretation. The table below summarizes several central quantities.
| Symbol or Term | Meaning | Typical Unit or Scale | Metabolic Interpretation |
|---|---|---|---|
| \(N(t)\) | Abundance at time \(t\) | cells, organisms, biomass proxy, or count | Population or culture size under metabolic conditions |
| \(N_0\) | Initial abundance | same as \(N(t)\) | Starting abundance for growth analysis |
| \(r\) | Per-capita growth rate | per unit time | Rate of abundance increase under specified conditions |
| \(K\) | Carrying capacity | same as \(N(t)\) | Approximate upper abundance supported by available resources and conditions |
| \(\mu(S)\) | Substrate-dependent growth rate | per unit time | Growth rate as a function of substrate availability |
| \(\mu_{\max}\) | Maximum growth rate | per unit time | Upper growth-rate limit under favorable substrate conditions |
| \(S\) | Substrate concentration or stoichiometric matrix, depending on context | concentration or matrix | Resource level in Monod modeling or reaction stoichiometry in flux balance |
| \(K_s\) | Half-saturation constant | same unit as substrate concentration | Substrate level associated with half of maximum growth rate |
| \(Y_{X/S}\) | Biomass yield on substrate | g/g, mol/mol, or biomass per substrate unit | Efficiency of substrate conversion into biomass |
| \(\Delta X\) | Change in biomass | g/L, cells, dry mass, or biomass unit | Biomass produced during a growth interval |
| \(\Delta S\) | Substrate consumed | g/L, mol/L, or substrate unit | Resource consumed during a growth interval |
| \(J\) | Flux or pathway throughput | amount per area per time, concentration per time, or model unit | Rate of transport or metabolic pathway flow |
| \(D\) | Diffusion coefficient | area per time | How rapidly a molecule spreads through a medium |
| \(C\) | Concentration | molarity, mass per volume, or relative abundance | Amount of nutrient, metabolite, oxygen, or product per volume |
| \(v_i\) | Step-specific capacity or reaction flux | rate or flux unit | Capacity of a pathway step or reaction |
| \(v\) | Flux vector | reaction fluxes | Reaction-rate vector in constraint-based metabolic modeling |
The table shows why metabolic analysis requires careful context. The same symbol may be used differently across modeling traditions. A growth rate, flux, yield, or substrate concentration becomes biologically meaningful only when linked to the organism, pathway, measurement method, and environmental conditions.
Worked Example: Growth Rate, Doubling Time, and Biomass Yield
Suppose a culture begins with \(N_0=1.0 \times 10^5\) cells and grows to \(4.0 \times 10^5\) cells in 48 hours during its early exponential phase. Under the exponential growth model:
Interpretation: Abundance changes according to initial abundance, growth rate, and elapsed time.
Substituting the values:
Interpretation: The observed fourfold increase can be used to estimate the growth rate.
Dividing both sides by \(1.0 \times 10^5\):
Interpretation: The culture increased fourfold during the observation interval.
Taking the natural logarithm:
Interpretation: The estimated growth rate is approximately 0.0289 per hour.
The doubling time is:
Interpretation: At this estimated rate, the culture doubles approximately every 24 hours.
This is useful because it converts raw abundance data into biologically interpretable parameters. Similar calculations are routine in cell culture, microbial growth, plankton studies, fermentation, treatment-response analysis, and ecological productivity work.
Biomass yield asks a different question. Suppose a microbial culture increases from 0.20 g/L to 0.95 g/L biomass while consuming 1.50 g/L glucose. Then:
Interpretation: Biomass increased by 0.75 grams per liter.
Interpretation: The culture consumed 1.50 grams per liter of substrate.
so:
Interpretation: The culture converted substrate into biomass with a yield of 0.50 g/g.
This is useful because yield captures a different biological question from growth rate alone: not how fast biomass accumulates, but how economically substrate is converted into biomass.
Computational Modeling
Computational modeling helps make metabolism explicit because metabolic life is dynamic, rate-dependent, and constrained. Growth curves estimate biological increase through time. Monod models represent nutrient limitation. Yield calculations summarize conversion efficiency. Allocation models distinguish growth, maintenance, product formation, and loss. Flux-balance models express the constraint that reactions must obey mass balance. Energy-budget models formalize tradeoffs among growth, repair, survival, and production.
The selected examples below focus on compact, reusable workflows: exponential growth fitting, doubling-time estimation, biomass yield, maintenance allocation, Monod substrate response, logistic growth under stress, and toy flux-balance analysis. The GitHub repository extends the same logic into richer workflows for oxygen consumption, ATP accounting, pathway bottleneck analysis, respirometry summaries, metabolic condition scoring, SQL provenance, notebooks, validation scripts, and multi-language scientific-computing examples.
The purpose is not to reduce metabolism to equations. The purpose is to make metabolic evidence inspectable. A metabolic claim becomes stronger when substrates, rates, yields, assumptions, units, model choices, and biological interpretation are documented together.
R Workflow: Growth, Yield, Allocation, and Monod Response
R is useful for metabolic analysis because it supports growth-curve fitting, yield summaries, allocation tables, statistical comparison, and reproducible reporting. The following workflow estimates exponential growth and doubling time, summarizes substrate allocation, calculates biomass yield, and models a Monod-style substrate-limited growth response.
# Metabolism, Energy, and Biological Function Workflow
#
# This workflow demonstrates four quantitative metabolism tasks:
#
# 1. Fit exponential growth and estimate doubling time.
# 2. Summarize substrate allocation across biomass, maintenance, product, and loss.
# 3. Calculate biomass yield on substrate.
# 4. Model Monod-style substrate-limited growth.
#
# These examples can be adapted for cell culture, microbial ecology,
# phytoplankton studies, fermentation, wastewater treatment,
# bioprocessing, environmental biotechnology, and metabolic engineering.
library(tibble)
library(dplyr)
# ------------------------------------------------------------
# 1. Fit exponential growth and estimate doubling time
# ------------------------------------------------------------
growth_data <- tibble(
time_h = c(0, 12, 24, 36, 48),
abundance = c(1.0e5, 1.4e5, 2.0e5, 2.8e5, 4.0e5)
)
growth_model <- lm(log(abundance) ~ time_h, data = growth_data)
growth_summary <- tibble(
growth_rate_per_h = coef(growth_model)[["time_h"]],
estimated_initial_abundance = exp(coef(growth_model)[["(Intercept)"]]),
doubling_time_h = log(2) / growth_rate_per_h,
r_squared_log_space = summary(growth_model)$r.squared
)
growth_fit <- growth_data %>%
mutate(
predicted_abundance = exp(predict(growth_model)),
residual = abundance - predicted_abundance
)
# ------------------------------------------------------------
# 2. Biomass yield and maintenance allocation
# ------------------------------------------------------------
substrate_input_g_L <- 2.0
allocation_df <- tibble(
pool = c("biomass", "maintenance", "product"),
substrate_g_L = c(0.90, 0.70, 0.25)
)
allocation_df <- allocation_df %>%
bind_rows(
tibble(
pool = "loss",
substrate_g_L = substrate_input_g_L - sum(allocation_df$substrate_g_L)
)
) %>%
mutate(
fraction_of_input = substrate_g_L / substrate_input_g_L
)
yield_summary <- tibble(
substrate_input_g_L = substrate_input_g_L,
biomass_generated_g_L = allocation_df$substrate_g_L[allocation_df$pool == "biomass"],
biomass_yield_g_g = biomass_generated_g_L / substrate_input_g_L,
maintenance_fraction = allocation_df$fraction_of_input[allocation_df$pool == "maintenance"],
product_fraction = allocation_df$fraction_of_input[allocation_df$pool == "product"]
)
# ------------------------------------------------------------
# 3. Monod-style substrate-limited growth
# ------------------------------------------------------------
monod_df <- tibble(
substrate_mM = seq(0, 20, length.out = 200)
) %>%
mutate(
mu_max_per_h = 0.08,
Ks_mM = 2.5,
growth_rate_per_h =
mu_max_per_h * substrate_mM / (Ks_mM + substrate_mM)
)
half_saturation_row <- monod_df %>%
slice(which.min(abs(growth_rate_per_h - unique(mu_max_per_h) / 2)))
print(round(growth_summary, 4))
print(round(growth_fit, 2))
print(round(allocation_df, 4))
print(round(yield_summary, 4))
print(head(round(monod_df, 4), 12))
print(tail(round(monod_df, 4), 12))
print(round(half_saturation_row, 4))This R workflow is useful because it produces directly interpretable quantities: growth rate, doubling time, yield, maintenance fraction, product allocation, and substrate-limited growth response. A working scientist could extend it to treatment comparisons, confidence intervals, nonlinear growth models, replicate-aware analyses, bioreactor time courses, or ecological productivity datasets.
Python Workflow: Logistic Growth, Allocation, and Toy Flux Balance
Python is useful for metabolism because it supports simulation, parameter sweeps, constraint logic, workflow automation, and reproducible computational analysis. The following workflow simulates logistic growth under metabolic stress, summarizes substrate allocation, and implements a transparent toy flux-balance analysis.
"""
Metabolism, Energy, and Biological Function Workflow
This workflow demonstrates three quantitative metabolism tasks:
1. Simulate logistic growth under control and stressed conditions.
2. Summarize substrate allocation and biomass yield.
3. Run a transparent toy flux-balance analysis.
The examples are compact, but the same structures can be extended to
cell culture, microbial ecology, phytoplankton studies, fermentation,
bioprocessing, wastewater treatment, environmental biotechnology,
metabolic engineering, and systems biology.
"""
from __future__ import annotations
import itertools
import numpy as np
import pandas as pd
def logistic_growth(
time: np.ndarray,
initial_abundance: float,
growth_rate: float,
carrying_capacity: float,
) -> np.ndarray:
"""
Calculate a logistic growth trajectory.
"""
if initial_abundance <= 0:
raise ValueError("Initial abundance must be positive.")
if carrying_capacity <= 0:
raise ValueError("Carrying capacity must be positive.")
if initial_abundance > carrying_capacity:
raise ValueError("Initial abundance should not exceed carrying capacity.")
return carrying_capacity / (
1.0
+ ((carrying_capacity - initial_abundance) / initial_abundance)
* np.exp(-growth_rate * time)
)
def simulate_growth_under_metabolic_stress() -> tuple[pd.DataFrame, pd.DataFrame]:
"""
Compare logistic growth under control and stressed conditions.
"""
time_h = np.linspace(0, 96, 400)
initial_abundance = 1.0e5
carrying_capacity = 1.0e6
scenarios = {
"control": 0.035,
"stressed": 0.020,
"strongly_limited": 0.012,
}
rows = []
for scenario, growth_rate in scenarios.items():
abundance = logistic_growth(
time=time_h,
initial_abundance=initial_abundance,
growth_rate=growth_rate,
carrying_capacity=carrying_capacity,
)
rows.append(
pd.DataFrame(
{
"scenario": scenario,
"time_h": time_h,
"abundance": abundance,
"growth_rate_per_h": growth_rate,
"initial_doubling_time_h": np.log(2.0) / growth_rate,
}
)
)
growth_df = pd.concat(rows, ignore_index=True)
summary_df = (
growth_df.groupby("scenario")
.agg(
growth_rate_per_h=("growth_rate_per_h", "first"),
initial_doubling_time_h=("initial_doubling_time_h", "first"),
final_abundance=("abundance", "last"),
)
.reset_index()
)
return growth_df, summary_df
def substrate_allocation_summary() -> tuple[pd.DataFrame, pd.DataFrame]:
"""
Summarize substrate allocation among biomass, maintenance, product, and loss.
"""
substrate_input = 2.0
allocation = {
"biomass": 0.90,
"maintenance": 0.70,
"product": 0.25,
}
allocation["loss"] = substrate_input - sum(allocation.values())
allocation_df = pd.DataFrame(
{
"pool": list(allocation.keys()),
"substrate_g_L": list(allocation.values()),
}
)
allocation_df["fraction_of_input"] = (
allocation_df["substrate_g_L"] / substrate_input
)
summary = pd.DataFrame(
{
"substrate_input_g_L": [substrate_input],
"biomass_yield_g_g": [allocation["biomass"] / substrate_input],
"maintenance_fraction": [allocation["maintenance"] / substrate_input],
"product_fraction": [allocation["product"] / substrate_input],
"loss_fraction": [allocation["loss"] / substrate_input],
}
)
return allocation_df, summary
def toy_flux_balance_analysis() -> pd.DataFrame:
"""
Run a transparent toy flux-balance analysis.
Toy steady-state system:
glucose uptake -> precursor
precursor -> biomass
precursor -> product
precursor -> respiration
This is not a genome-scale model. It is a simple teaching scaffold
showing that feasible fluxes must satisfy balance constraints and
that different objectives imply different allocations.
"""
glucose_uptake = 10.0
candidate_fluxes = []
for biomass_flux, product_flux, respiration_flux in itertools.product(
np.linspace(0, 10, 101),
np.linspace(0, 10, 101),
np.linspace(0, 10, 101),
):
precursor_balance = (
glucose_uptake - biomass_flux - product_flux - respiration_flux
)
if abs(precursor_balance) <= 0.05:
objective = biomass_flux + 0.25 * product_flux
candidate_fluxes.append(
{
"glucose_uptake": glucose_uptake,
"biomass_flux": biomass_flux,
"product_flux": product_flux,
"respiration_flux": respiration_flux,
"precursor_balance": precursor_balance,
"objective": objective,
}
)
flux_df = pd.DataFrame(candidate_fluxes)
return flux_df.sort_values("objective", ascending=False).head(10)
def main() -> None:
"""
Run compact metabolism workflows.
"""
growth_df, growth_summary = simulate_growth_under_metabolic_stress()
allocation_df, allocation_summary = substrate_allocation_summary()
best_fluxes = toy_flux_balance_analysis()
print("Logistic growth under metabolic stress:")
print(growth_df.head(12).round(3).to_string(index=False))
print(growth_df.tail(12).round(3).to_string(index=False))
print("\nGrowth scenario summary:")
print(growth_summary.round(4).to_string(index=False))
print("\nSubstrate allocation:")
print(allocation_df.round(4).to_string(index=False))
print("\nAllocation summary:")
print(allocation_summary.round(4).to_string(index=False))
print("\nToy flux-balance analysis: top feasible allocations")
print(best_fluxes.round(4).to_string(index=False))
if __name__ == "__main__":
main()This Python workflow is useful because it distinguishes three forms of metabolic reasoning: growth under constraint, substrate allocation among competing demands, and flux feasibility under mass-balance logic. The examples are simplified, but they provide transparent scaffolding for richer metabolic engineering, microbial ecology, cell-culture, and environmental biotechnology workflows.
GitHub Repository
The article body includes compact R and Python examples so the biological and scientific argument remains readable. The full repository expands those examples into a more rigorous computational metabolism workflow, including growth-curve fitting, logistic and Monod growth models, biomass-yield estimation, maintenance allocation, oxygen-consumption summaries, energy budgets, simplified ATP accounting, pathway bottleneck analysis, toy flux-balance analysis, metabolic condition scoring, SQL provenance structures, validation notes, reproducible data files, and full-stack scientific-computing examples across Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, and notebooks.
Complete Code Repository
The full code distribution for this article, including selected article examples, expanded computational workflows, reproducible data structures, provenance documentation, validation notes, and full-stack scientific-computing scaffolding, is available on GitHub.
Limits, Scaling, and Modern Metabolic Thinking
Metabolism is foundational, but it is not simple. Metabolic pathways are context-dependent, tissue-specific, developmentally regulated, environmentally constrained, and often nonlinear. A result measured in a cell line may not map cleanly onto a whole organism. An organismal metabolic rate may not explain ecosystem-level nutrient cycling without ecological context. A microbial process observed in a flask may differ substantially in soils, sediments, estuaries, biofilms, hosts, or bioreactors.
This is why modern metabolic thinking increasingly emphasizes scale and integration. Metabolism must be understood in cells, but also in tissues, organisms, populations, microbiomes, ecosystems, and planetary cycles. It must be linked to physiology, ecology, evolution, environmental constraint, and systems biology. Quantitative models help, but they do not eliminate the need for careful biological judgment.
Models and workflows are useful because they clarify assumptions, expose bottlenecks, and make comparisons possible. But a growth curve is not a complete metabolic theory, a yield coefficient is not a full energy budget, and a toy flux-balance model is not a genome-scale reconstruction. Quantitative metabolism is strongest when it supports biological interpretation rather than replacing it.
In that sense, metabolism is a model case for modern biology itself: mechanistic, quantitative, historically shaped, environmentally embedded, computationally analyzable, and irreducible to any one level of explanation. The strongest metabolic analysis connects pathway detail with ecological and physiological context.
Why This Matters for Scientific Work
For working scientists, metabolism matters because many biological problems are best understood as constrained throughput problems rather than as static descriptions of molecules or traits. A microbial bloom, plant stress response, animal growth trajectory, clinical fatigue syndrome, tumor microenvironment, forest carbon sink, or wastewater process may all depend on rates of uptake, energetic allocation, oxygen limitation, redox balance, or pathway bottlenecks rather than on one isolated variable.
This means metabolism should often be treated not as a background chapter in biology, but as one of the major frameworks through which biological systems become analytically intelligible. Ecologists need it because trophic dynamics and nutrient cycles are metabolically grounded. Physiologists need it because function depends on controlled energy flow. Microbiologists need it because microbial life is often best distinguished by energetic and redox strategy. Biomedical scientists need it because disease frequently involves metabolic reprogramming, insufficiency, or misregulation. Computational biologists need it because metabolism is one of the clearest networked, rate-dependent, multiscale problems in the life sciences.
The scientific importance of metabolism lies partly in this breadth. It is one of the principal ways biology explains how living systems remain active, resilient, vulnerable, and possible under real conditions of energetic constraint.
Metabolism is also practically actionable. Substrate levels can be measured. Growth rates can be fitted. Oxygen consumption can be tracked. Fluxes can be modeled. Biomass yields can be estimated. Energy allocation can be compared across conditions. These measurements connect biological explanation to experimental design, medical interpretation, ecological monitoring, and biotechnological optimization.
Conclusion
Metabolism, energy, and biological function show that life depends on organized transformations of matter and energy. Metabolism powers maintenance, growth, reproduction, regulation, repair, and response. It links ATP and redox chemistry to physiology, ecological productivity, microbial cycling, plant growth, disease processes, biotechnology, and the broader circulation of energy and matter across living systems.
To understand metabolism is therefore to understand one of the deepest conditions of life itself. Living systems persist not because they escape material constraint, but because they regulate energy and matter with extraordinary precision under changing conditions. That is why metabolism remains central not only to cell biology and physiology, but also to ecology, marine and freshwater science, soil biology, plant science, microbiology, agroecology, forestry, disease ecology, biotechnology, and systems biology.
Metabolism is thus more than a topic within biology. It is one of the principal ways biology understands how life remains active, resilient, vulnerable, and possible. Modern quantitative and computational workflows deepen that understanding by making growth, yield, flux, allocation, and energetic constraint more transparent. The result is a biology that can connect molecular reactions to organisms, ecosystems, medicine, and sustainable systems of life.
Related Articles
- Biology
- Cell Theory and the Basic Unit of Life
- Biomolecules and the Chemical Basis of Life
- Water, Energy, and the Material Conditions of Life
- Cell Structure, Membranes, and Organelles
- Enzymes, Regulation, and Biochemical Pathways
- Molecular Biology and the Flow of Genetic Information
- Cell Signaling, Communication, and Biological Coordination
- Physiology and the Regulation of Living Systems
- Microbiology and the Hidden Majority of Life
- Biogeochemical Cycles and the Conditions of Habitability
- Plant Biology and the Life of Primary Producers
- Fungi and the Networks of Decomposition and Exchange
- Ecology and the Interdependence of Life
- Systems Biology and the Logic of Biological Integration
Further Reading
- Alberts, B. et al. (2002) ‘How cells obtain energy from food’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26882/
- Alberts, B. et al. (2002) ‘Metabolic energy’, in The Cell: A Molecular Approach. 2nd edn. Sunderland, MA: Sinauer Associates. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9903/
- Alberts, B. et al. (2002) ‘The mitochondrion’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26894/
- Berg, J.M., Tymoczko, J.L. and Stryer, L. (2002) Biochemistry. 5th edn. New York: W.H. Freeman. Available at: https://www.ncbi.nlm.nih.gov/books/NBK21154/
- de Nava, A.S.L. et al. (2022) ‘Physiology, metabolism’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK546690/
- Deshpande, O.A. et al. (2023) ‘Biochemistry, oxidative phosphorylation’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK553192/
- Dunn, J. and Grider, M.H. (2023) ‘Physiology, adenosine triphosphate’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK553175/
- Monod, J. (1949) ‘The growth of bacterial cultures’, Annual Review of Microbiology, 3, pp. 371–394. Available at: https://www.annualreviews.org/content/journals/10.1146/annurev.mi.03.100149.002103
- Orth, J.D., Thiele, I. and Palsson, B.Ø. (2010) ‘What is flux balance analysis?’, Nature Biotechnology, 28, pp. 245–248. Available at: https://www.nature.com/articles/nbt.1614
References
- Alberts, B. et al. (2002) ‘How cells obtain energy from food’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26882/
- Alberts, B. et al. (2002) ‘Metabolic energy’, in The Cell: A Molecular Approach. 2nd edn. Sunderland, MA: Sinauer Associates. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9903/
- Alberts, B. et al. (2002) ‘The mitochondrion’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26894/
- Berg, J.M., Tymoczko, J.L. and Stryer, L. (2002) Biochemistry. 5th edn. New York: W.H. Freeman. Available at: https://www.ncbi.nlm.nih.gov/books/NBK21154/
- de Nava, A.S.L. et al. (2022) ‘Physiology, metabolism’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK546690/
- Deshpande, O.A. et al. (2023) ‘Biochemistry, oxidative phosphorylation’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK553192/
- Dunn, J. and Grider, M.H. (2023) ‘Physiology, adenosine triphosphate’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK553175/
- Monod, J. (1949) ‘The growth of bacterial cultures’, Annual Review of Microbiology, 3, pp. 371–394. Available at: https://www.annualreviews.org/content/journals/10.1146/annurev.mi.03.100149.002103
- Orth, J.D., Thiele, I. and Palsson, B.Ø. (2010) ‘What is flux balance analysis?’, Nature Biotechnology, 28, pp. 245–248. Available at: https://www.nature.com/articles/nbt.1614