
Last Updated May 28, 2026
Development, differentiation, and the making of organisms examine how living systems progress from relatively simple beginnings to complex, organized, multi-scale forms through regulated processes of cell division, pattern formation, specification, morphogenesis, growth, and functional specialization. Development is one of the central problems of biology because organisms are not assembled from preexisting miniature parts. They are produced through dynamic, highly coordinated processes in which cells divide, acquire distinct identities, communicate with one another, move through tissues, respond to positional information, and collectively generate organs, body plans, life-history stages, and organismal function.
This article develops Development, Differentiation, and the Making of Organisms as a foundational article within the Biology knowledge series. It treats developmental biology not as a narrow embryological specialty, but as one of biology’s major organizing frameworks: the field that explains how heredity becomes form, how regulation becomes tissue organization, how cells become specialized, how organisms coordinate life cycles, and how environmental conditions shape biological possibility during sensitive windows of growth and transformation.
Main Library
Publications
Article Map
Biology
Related Topic
Chemistry
Related Topic
Earth Science
Related Topic
Environmental Science
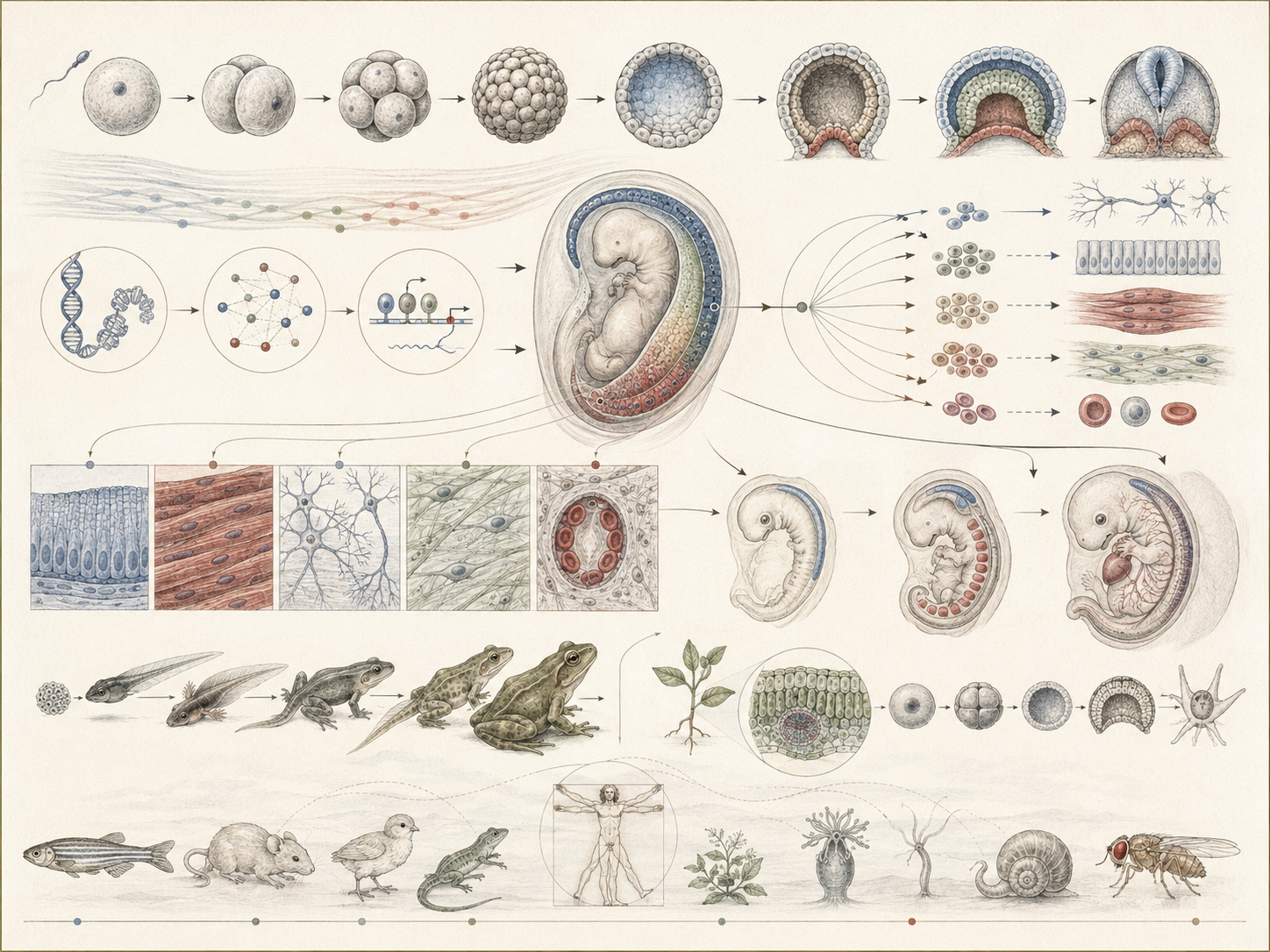
The article develops development, differentiation, and organism formation across cell fate, positional information, morphogen gradients, gene expression, epigenetic regulation, cell signaling, morphogenesis, tissue mechanics, growth, organ formation, life-cycle coordination, plant and animal development, regeneration, stem cells, disease, ecological stress, evolutionary developmental biology, biotechnology, and computational developmental systems.
The article is written for developmental biologists, cell biologists, evolutionary biologists, plant scientists, animal biologists, marine and freshwater researchers, biomedical scientists, stem-cell researchers, regenerative-medicine readers, disease ecologists, conservation practitioners, restoration ecologists, systems biologists, and computational biology readers who need a rigorous account of how organisms are made through growth, differentiation, patterning, regulation, and environmental response.
The article also extends developmental biology into quantitative and computational analysis through exponential growth, logistic growth, doubling-time estimation, branching differentiation dynamics, morphogen-gradient thresholding, reaction-diffusion patterning, stage-dependent growth, Markov-style developmental state transitions, R workflows, Python workflows, SQL provenance structures, and a linked full-stack GitHub repository containing Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, notebooks, data files, validation notes, and reproducibility documentation.
What development and differentiation are
Development is the set of biological processes through which organisms progress across life stages and generate organized form, structure, and function. Differentiation is the process through which cells become specialized in identity, behavior, and role, even when they descend from genetically similar or genetically identical predecessors. Together, development and differentiation explain how living systems become structured rather than remaining undifferentiated collections of cells, tissues, or biological potential.
This makes developmental biology one of the deepest branches of biology. It asks how bodies are made, how tissues acquire identity, how cells interpret position, how growth is coordinated with form, how life cycles proceed, how organs emerge, and how similar genomes can support radically different cell types and anatomical outcomes. Organisms are therefore not simply inherited as finished entities. They are made through regulated, time-dependent biological processes.
Development also links continuity and novelty. Organisms preserve hereditary information across generations, but each new organism must still be built through the unfolding of developmental processes. Development is therefore the bridge between heredity and embodied form. In a deeper sense, development is where information becomes geometry, where regulation becomes anatomy, and where the potential of a genome becomes the realized architecture of an organism.
Development is also not confined to embryos. Plants continue developmental growth through meristems. Fungi shift among growth forms. Microbes can differentiate into specialized states, spores, or biofilms. Animals pass through juvenile stages, metamorphosis, regeneration, maturation, and aging. Development is therefore a general biological problem: how living systems organize change across time.
Development as a central problem of biology
Development is a central biological problem because living form is produced, not merely given. An embryo, seed, spore, larva, meristem, or stem-cell population does not contain a fully realized adult structure in miniature. Instead, development proceeds through regulated events such as cleavage, growth, specification, migration, adhesion, patterning, differentiation, apoptosis, morphogenesis, and tissue remodeling. This is why developmental biology became historically foundational: it gave biology a way to understand how organized complexity emerges through time.
This problem is challenging because development requires coordination across scales. Molecular regulation influences cell behavior. Cell behavior shapes tissues. Tissues generate organs. Organs contribute to organismal identity and life-history function. The making of organisms therefore cannot be reduced to one level alone. It must be understood across nested systems of causation, constraint, and feedback.
Development is also central because it is one of the clearest places where biology becomes explicitly temporal. Organisms are not static objects but unfolding processes, and developmental biology studies the logic of that unfolding. Timing, sequence, threshold behavior, irreversible commitment, reversible plasticity, and spatial coordination all matter. A developmental process succeeds not merely because all the right components exist, but because they are activated in the right order, at the right scale, in the right place, and under the right conditions.
This makes development a model case for systems biology. It is dynamic, multi-scale, spatially organized, historically constrained, and environmentally responsive. It shows why biology cannot be understood only through inventories of molecules or genes. The order in which processes occur, the relationships among components, and the spatial organization of cells and tissues are all part of the explanation.
Cell fate specification and positional information
One of development’s key problems is how cells acquire distinct fates. Early in development, cells may be relatively similar, yet later they become muscle, nerve, epidermis, vascular tissue, gut lining, reproductive tissue, root hair, pollen grain, immune cell, endocrine cell, or countless other specialized forms. Developmental biology explains this through specification, induction, signaling, lineage history, gene regulation, epigenetic state, and positional information. Cells often respond not only to lineage, but also to where they are, what neighboring cells signal, and what gradients or local environments they encounter.
The concept of positional information is especially important. A cell can adopt a fate partly because it occupies a particular position within a developing field. Morphogens are often used to explain this logic. A morphogen is a signaling molecule whose concentration can help specify different cell fates across a spatial field. Cells exposed to high, intermediate, or low concentrations may activate different regulatory programs. Development therefore becomes intelligible as both a spatial and temporal process.
Cell fate is therefore not usually a matter of intrinsic essence alone. It emerges through regulated interaction among genomes, signals, location, mechanics, and developmental history. This is one of the central lessons of developmental biology. Modern work expands this further by showing that cells often integrate multiple signals simultaneously, combine lineage memory with local information, and transition through intermediate competence states before reaching more stable identities.
This has major implications beyond embryology. Stem-cell biology, regenerative medicine, cancer biology, plant development, microbial differentiation, organoid modeling, and tissue engineering all depend on understanding how cell fate is specified, stabilized, reversed, or misdirected. A cell type is not simply a label. It is a regulated state produced through developmental history.
Differentiation and functional specialization
Differentiation is the process by which cells become specialized in form and function. The resulting differences can be profound: one cell type may become contractile, another electrically excitable, another secretory, another photosynthetic, another immune-responsive, another absorptive, another structural, another reproductive, and another stem-like or proliferative. Yet these outcomes often arise from shared genetic material interpreted through different regulatory states and developmental contexts.
This makes differentiation one of the clearest demonstrations that biological identity depends on regulation and context rather than sequence alone. The same genome can support many different cell types because gene expression is selective, regulated, and historically patterned through development. Differentiation is therefore one of the strongest arguments against simplistic genetic determinism. A genome provides inherited potential, but development organizes that potential into specific cell states, tissue roles, and organismal functions.
Functional specialization also matters because organismal complexity depends on division of labor. Multicellularity becomes biologically meaningful when cells do different things in coordinated ways. Differentiation is one of the main routes by which this becomes possible. In quantitative terms, development often involves progressive restriction of possible cell states together with progressive gain of specialized function, a transition that can be analyzed as both regulatory commitment and state-space narrowing.
Differentiation is also reversible or plastic in certain contexts. Some cells can dedifferentiate, transdifferentiate, or regain stem-like features under specific conditions. This plasticity matters in regeneration, wound repair, cancer, immune response, plant tissue culture, and biotechnology. Developmental biology therefore studies not only how cells become specialized, but also how specialization is maintained, modified, disrupted, or experimentally redirected.
Morphogenesis and the making of form
Morphogenesis refers to the processes through which tissues, organs, and body structures acquire shape and arrangement. Development is not only about producing the right cell types. It is also about organizing them into the right spatial relationships. A body is more than a collection of differentiated cells. It is a structured system in which cells must adhere, migrate, proliferate, die, fold, branch, fuse, polarize, and rearrange in coordinated ways.
This is why cell adhesion, epithelial-mesenchymal transitions, branching, tissue folding, matrix remodeling, polarity, mechanical force, and structural pattern formation are so important in developmental biology. Morphogenesis explains how order becomes visible and anatomical. A retina, leaf, limb, root system, vascular network, branching lung, fungal mycelium, epithelial fold, or floral structure is not produced simply by adding more cells. It is produced through dynamic organization.
The making of form therefore reveals that development is both chemical and mechanical, both genetic and spatial. Organismal structure emerges through coordinated interactions among signaling, cell behavior, tissue geometry, extracellular matrix, and physical forces. Contemporary developmental biology increasingly treats morphogenesis as a problem of collective dynamics, force transmission, polarity, curvature generation, branching logic, and local rule-based organization rather than only a list of molecular components.
This also explains why developmental disruption can be systemic. If cells differentiate but fail to organize, function may still fail. If growth occurs without proper patterning, tissues may become malformed. If tissue mechanics are altered, signaling may be misread. Morphogenesis connects microscopic cellular behavior to macroscopic biological form.
Growth, patterning, and organismal organization
Growth is an obvious part of development, but growth alone does not explain organismal organization. Patterning is equally important because tissues and structures must arise in the right positions, proportions, and relationships. Growth without pattern would produce mass without coherent form. Pattern without growth would remain unrealized. Development requires both.
Patterning includes axial organization, segmentation, symmetry, polarity, repeated structures, branching geometry, organ primordia, differentiated tissue zones, and spatial boundaries. It explains how organisms come to have fronts and backs, roots and shoots, dorsal and ventral domains, branched organs, repeated units, or differentiated tissue regions. These are among the main ways developmental systems generate organismal coherence.
This means the making of organisms is not only a matter of producing cells or organs in isolation. It is a matter of producing ordered relations among them. Pattern is therefore one of development’s deepest themes. In advanced developmental reasoning, pattern is often treated as an emergent property of interacting gradients, feedback loops, mechanical constraints, reaction-diffusion processes, lineage-dependent competence fields, and local tissue interactions.
Growth and patterning also create problems of scaling. As organisms grow, pattern must remain proportionate enough to produce viable function. Different species, organs, and life stages solve this problem in different ways. Development therefore connects size, proportion, timing, and geometry. It is the science of regulated biological construction across changing scale.
Developmental regulation and the coordination of life cycles
Development does not stop at embryogenesis. Many organisms undergo life-history transitions that include larval stages, metamorphosis, dormancy, germination, flowering, reproduction, regeneration, senescence, seasonal developmental shifts, or environmentally triggered developmental pathways. Developmental regulation therefore helps coordinate life cycles across ecological and physiological time.
This is important because development is not always a one-directional march toward a fixed adult form. In many organisms, development includes branching pathways, environmentally responsive timing, arrested states, plastic transitions, or repeated cycles of growth and differentiation. Seeds, spores, insect metamorphosis, amphibian transformation, plant meristem activity, fungal life cycles, colonial systems, and regenerative systems all show that development can be flexible, staged, and conditional.
Biology is therefore strongest when it understands development as a regulated life-cycle process rather than only as early embryonic change. The making of organisms often extends across the entire life history. In many systems, developmental timing is one of the decisive variables through which ecology enters biology, because shifts in seasonality, temperature, hydrology, nutrient availability, disease exposure, or disturbance can reconfigure developmental schedules and thereby alter survival and reproduction.
This makes development highly relevant to climate change, conservation, agriculture, disease ecology, and restoration biology. If environmental conditions shift during sensitive developmental windows, the effects may appear later as altered body size, reproductive success, immune function, stress tolerance, or population recruitment. Development is one of the ways ecological pressure becomes embodied biological outcome.
Development in physiology, function, and organismal identity
Development matters because biological function depends on it. Organs do not simply appear fully formed. They are constructed through developmental processes that establish cell types, tissue interactions, structural arrangement, vascular organization, innervation, polarity, patterning, and regulatory capacity. Physiology therefore depends on development because function requires correctly produced form.
Organismal identity also depends on development. A species’ life form, anatomy, physiology, behavior, and functional capacities are expressed through the developmental pathways by which bodies are made. Development is therefore one of the major ways heredity becomes embodied. What is inherited is not merely a trait list, but the capacity to generate organized living structure through regulated change.
This is why developmental biology belongs at the intersection of genetics, molecular biology, cell biology, physiology, morphology, ecology, evolution, and systems biology. It explains how biological possibility becomes realized form. It also explains why developmental disruption can cascade so powerfully: failures of timing, signaling, cell migration, polarity, tissue mechanics, or cell-state control often produce systemic consequences because function depends on correctly assembled organization across many scales at once.
Development also clarifies why adult traits cannot always be understood from adult conditions alone. Some vulnerabilities, capacities, or physiological differences originate in earlier developmental contexts. Early nutrition, environmental stress, endocrine exposure, temperature, oxygen, disease, or toxicant exposure can alter later biological performance. Developmental biology therefore links life history to physiology and ecology across time.
Ecology, evolution, and sustainability-adjacent biology
Development is deeply relevant to ecology and sustainability-adjacent biology because organisms encounter environments developmentally, not only as fully formed adults. Temperature, moisture, salinity, nutrients, toxins, crowding, predation risk, disease exposure, hypoxia, acidity, light, and other ecological factors can shape developmental outcomes, timing, survivorship, and plasticity. Developmental response is therefore one of the ways environmental change becomes biologically consequential.
Evolutionary biology is equally important here because developmental systems influence what kinds of variation can be expressed, tolerated, and selected. Evolutionary developmental biology emerged precisely because development shapes the routes by which novelty and form can evolve. Development is therefore not only a downstream expression of genes. It is part of the architecture through which evolutionary possibility is filtered.
This matters for sustainability because populations and ecosystems under stress do not respond only through adult survival or mortality. They also respond through altered recruitment, disrupted life cycles, developmental instability, plastic adjustment, failed metamorphosis, delayed reproduction, impaired germination, and failures of differentiation or pattern formation. Development is thus part of resilience and vulnerability alike.
In restoration and conservation, this means recruitment bottlenecks, larval survival windows, germination cues, juvenile developmental transitions, seedling establishment, regeneration thresholds, and reproductive timing are often as important as adult abundance. A population may contain adults yet fail to sustain itself if developmental stages are disrupted. Sustainability-oriented biology must therefore treat development as part of ecological continuity.
Marine, freshwater, soil, plant, and microbial relevance
Marine biology makes developmental thinking especially important because many marine organisms pass through distinct developmental stages, including embryos, larvae, settlement phases, metamorphic transitions, and adult forms that may occupy very different ecological niches. Temperature, acidity, oxygen, salinity, pollutants, and food availability can all affect marine development in ways that shape population persistence and ecosystem structure.
Freshwater biology presents similar developmental stakes in amphibians, fishes, aquatic plants, invertebrates, and microbial communities responding to hydrologic disturbance, pollution, eutrophication, warming, fragmentation, and oxygen stress. In many freshwater systems, recruitment and early life stages are highly sensitive to environmental disruption. Developmental failure may appear later as population decline.
Plant science is equally developmental because roots, shoots, leaves, flowers, seeds, meristems, vascular systems, reproductive structures, and stress-response architectures all arise through ongoing developmental organization. Unlike many animals, plants retain strong developmental plasticity throughout life through meristems, branching, root growth, leaf production, flowering transitions, and environmental response.
Soil biology and microbiology also intersect with development, especially where spores, biofilms, differentiation states, symbiotic structures, fungal morphologies, and plant-microbe interactions shape ecological function. Agroecology, forestry, and restoration ecology therefore depend strongly on developmental understanding because establishment, growth, regeneration, and long-term resilience all begin with how organisms develop under real environmental conditions. Across these domains, life-history transitions are often ecologically decisive and highly vulnerable to environmental disruption.
Medical, biomedical, and disease ecology relevance
Development is foundational to medicine and biomedicine because many diseases arise through altered developmental processes, failed differentiation, abnormal morphogenesis, disrupted tissue organization, or misregulated signaling. Embryology, teratology, regenerative medicine, stem cell biology, oncology, developmental pathology, organoid modeling, tissue engineering, and congenital disease all depend on understanding how normal development proceeds and how it can fail.
Differentiation is especially important in cancer biology, where dedifferentiation, abnormal cell-state control, and disrupted developmental signaling can contribute to malignancy. Developmental defects can also arise through genetic change, environmental exposure, metabolic disturbance, endocrine disruption, infection, nutrient deficiency, or signaling disruption during sensitive periods. Development therefore connects heredity, environment, and pathology in especially direct ways.
Disease ecology expands this further because environmental disruption can alter developmental success across populations and species. Developmental sensitivity is therefore not only a medical issue. It is also an ecological one. Toxic exposures, endocrine disruption, nutrient stress, pathogen challenge, hypoxia, warming, and thermal anomalies can change developmental outcomes at both individual and population scales.
This perspective also matters for public health. Some biological outcomes are shaped by early developmental windows long before symptoms appear. Developmental biology therefore helps explain why prevention, environmental protection, maternal health, nutrition, exposure reduction, and early-life conditions can have long-term biological consequences. Development is not merely a beginning; it is a foundation.
Biotechnology, stem cells, and computational relevance
Biotechnology extends developmental biology into applied systems of stem cell research, tissue engineering, regenerative medicine, organoid modeling, agricultural improvement, developmental screening, and synthetic morphogenesis. Stem cells are especially important because they combine self-renewal with the capacity to differentiate into specialized cell types, making them central to both developmental biology and applied biomedical research. Developmental systems become experimentally tractable in these contexts because differentiation can be observed, induced, perturbed, and analyzed under controlled conditions.
Computational biology is increasingly important here because development involves branching trajectories, time-dependent changes, multiscale patterning, spatial fields, tissue mechanics, cell-state transitions, and large datasets from imaging, transcriptomics, epigenomics, and lineage tracing. Development is therefore one of the clearest cases where dynamic biological systems require quantitative and computational interpretation to become fully legible.
This makes developmental biology a strong bridge between basic biological theory and applied scientific practice. The same principles that explain how organisms are made can inform how tissues are modeled, repaired, engineered, regenerated, or compared under experimental conditions. Computationally, development is often treated through dynamical systems, state transitions, pseudotime trajectories, image-based morphometrics, reaction-diffusion models, agent-based simulations, branching-process inference, and spatial data analysis.
Developmental biotechnology also requires careful interpretation. Organoids are not full organs. Stem-cell differentiation in vitro is not identical to development in vivo. A computational trajectory is not proof of causal lineage. Developmental evidence becomes strongest when experimental perturbation, lineage tracing, spatial context, molecular state, and functional validation reinforce one another.
Quantitative developmental biology: mathematics, R, and Python
Development is not only descriptive. It is also quantitative. Growth rates, branching patterns, differentiation trajectories, spatial gradients, developmental timing, tissue expansion, and pattern formation can all be modeled mathematically and explored computationally. This does not reduce development to equations, but it makes developmental dynamics more explicit, testable, and reproducible.
A simple early growth approximation for a proliferating cell population is:
Interpretation: Cell number increases exponentially when growth proceeds at an approximately constant proportional rate.
where \(N(t)\) is the cell number at time \(t\), \(N_0\) is the initial cell number, and \(r\) is the growth rate. This is useful for early developmental proliferation or expanding cell populations under relatively unconstrained conditions.
The associated doubling time is:
Interpretation: Doubling time converts a fitted growth rate into the time required for the population to double.
This is useful because developmental growth is often easier to interpret as a timescale than as a raw exponential parameter.
Where developmental growth is limited by space, nutrients, tissue constraints, or regulatory feedback, a logistic model may be more realistic:
Interpretation: Logistic growth represents expansion under a constraint or carrying-capacity-like limit.
where \(K\) is a constraint parameter. This is useful because development often involves regulated growth under spatial, nutritional, mechanical, or signaling limits rather than unconstrained expansion.
A simple progenitor-to-differentiated transition can be represented as:
Interpretation: The progenitor compartment declines as cells leave that state.
Interpretation: The differentiated compartment gains cells at the rate progenitors transition.
where \(P\) is the progenitor compartment, \(D\) is the differentiated compartment, and \(k\) is the transition rate.
A branching model allows progenitors to feed two differentiated lineages:
Interpretation: Progenitors are depleted by two competing differentiation routes.
Interpretation: The first differentiated lineage gains cells according to transition rate \(k_1\).
Interpretation: The second differentiated lineage gains cells according to transition rate \(k_2\).
This is useful because real development often branches rather than moving through one linear progenitor-to-terminal path.
For spatial pattern formation, an activator-inhibitor style reaction-diffusion system can be expressed as:
Interpretation: The variable \(u\) changes through diffusion and local reaction dynamics.
Interpretation: The variable \(v\) changes through its own diffusion and coupled local reaction dynamics.
where \(u\) and \(v\) are interacting morphogen-like variables, \(D_u\) and \(D_v\) are diffusion terms, and \(f\) and \(g\) define local reaction kinetics. This matters because developmental patterning often depends on coupled local activation, inhibition, diffusion, and redistribution.
Variables, units, and developmental interpretation
Quantitative developmental biology depends on variables that connect growth, differentiation, pattern formation, state transitions, and biological interpretation. The table below summarizes several central quantities.
| Symbol or Term | Meaning | Typical Unit or Scale | Developmental Interpretation |
|---|---|---|---|
| \(N(t)\) | Cell number or population size at time \(t\) | cells, cell density, biomass, or normalized abundance | Size of a developing cell population or tissue compartment |
| \(N_0\) | Initial cell number | cells or normalized units | Starting size of a developmental population |
| \(r\) | Growth rate | per unit time | Rate of exponential or logistic expansion |
| \(t_d\) | Doubling time | time | Time required for a population to double under a fitted growth rate |
| \(K\) | Constraint or carrying-capacity-like parameter | cells, biomass, tissue size, or model-specific units | Upper limit or regulating constraint on growth |
| \(P\) | Progenitor compartment | cells, fraction, or normalized state abundance | Developmental state capable of generating differentiated descendants |
| \(D\) | Differentiated compartment | cells, fraction, or normalized state abundance | Population of cells that has transitioned into a specialized state |
| \(D_1, D_2\) | Differentiated lineages | cells, fraction, or normalized state abundance | Alternative developmental outputs from a shared progenitor pool |
| \(k\) | Differentiation transition rate | per unit time | Rate at which progenitors transition into a differentiated state |
| \(k_1, k_2\) | Branch-specific transition rates | per unit time | Relative allocation of progenitors into alternative lineages |
| \(u, v\) | Patterning or morphogen-like variables | concentration, signal intensity, or normalized field value | Spatially distributed variables that influence developmental patterning |
| \(D_u, D_v\) | Diffusion coefficients | area per time or model-specific spatial units per time | How quickly patterning variables spread across a spatial field |
| \(f(u,v), g(u,v)\) | Local reaction terms | concentration per time or model-specific units | Local production, decay, activation, or inhibition dynamics |
| Threshold | Boundary value for fate assignment | signal intensity or normalized concentration | Level at which a developmental signal produces a different cell fate |
| Transition matrix | Probabilities of moving between developmental states | probability from 0 to 1 | State-space model of developmental progression or cell-state composition |
The table shows why developmental quantities require context. A growth rate, differentiation rate, morphogen threshold, or state-transition probability becomes biologically meaningful only when linked to organism, tissue, developmental stage, measurement method, environmental condition, and model assumption.
Worked example: growth rate, doubling time, and lineage allocation
Suppose a developmental cell population begins with \(N_0=1.0\times10^4\) cells and reaches \(4.0\times10^4\) cells in 24 hours during an early proliferative phase. Under the exponential growth model:
Interpretation: Cell number grows according to initial cell number, growth rate, and elapsed time.
Substituting:
Interpretation: The cell population has increased fourfold over 24 hours.
Dividing both sides:
Interpretation: The unknown growth rate can be isolated through logarithms.
Taking the natural logarithm:
Interpretation: Log transformation turns exponential growth into a linear relation.
Solving:
Interpretation: The estimated proportional growth rate is approximately 0.0578 per hour.
The doubling time is:
Interpretation: The cell population doubles approximately every 12 hours under this simplified early-growth model.
This is useful because it turns developmental growth into an interpretable dynamic parameter rather than a purely descriptive observation.
Now suppose a progenitor pool has \(P=1000\) cells and two possible differentiation routes with rates \(k_1=0.14\) and \(k_2=0.09\) per time unit. At the starting moment, the instantaneous flux into lineage 1 is:
Interpretation: The first lineage initially receives 140 cells per time unit under this simplified model.
The instantaneous flux into lineage 2 is:
Interpretation: The second lineage initially receives 90 cells per time unit.
The progenitor pool declines at:
Interpretation: The progenitor compartment initially loses 230 cells per time unit across both differentiation routes.
This is useful because it shows how branching differentiation can be represented as allocation among competing developmental outcomes.
R and Python workflows
Computational modeling helps make developmental biology explicit because development is dynamic, spatial, branching, and stage-dependent. Growth models estimate proliferation. Logistic models represent constraints. Differentiation models track changes in cell-state composition. Morphogen-gradient thresholds formalize positional information. Reaction-diffusion models explore spatial self-organization. Stage-dependent growth models capture regime shifts. Markov-style transition matrices provide a simplified way to represent changing developmental-state probabilities.
The selected examples below focus on compact, reusable workflows: exponential growth fitting, doubling-time estimation, logistic growth, branching differentiation, morphogen thresholding, reaction-diffusion patterning, stage-dependent growth, and developmental state-transition modeling. The GitHub repository extends the same logic into richer workflows for SQL provenance, reproducible data files, validation notes, notebooks, organoid-style condition scoring, and multi-language scientific-computing examples.
The purpose is not to reduce development to code. The purpose is to make developmental reasoning inspectable. A developmental claim becomes stronger when growth assumptions, lineage-state definitions, spatial measurements, environmental conditions, stage timing, and analytical code are documented together.
R workflow: growth, logistic constraint, lineage splitting, and morphogen thresholds
R is useful for developmental biology because it supports growth modeling, tabular summaries, statistical fitting, differentiation-state analysis, and reproducible reporting. The following workflow estimates developmental growth rate and doubling time, simulates constrained logistic growth, models branching differentiation into two lineages, and assigns cell fates from a morphogen-gradient threshold model.
# Development, Differentiation, and the Making of Organisms Workflow
#
# This workflow demonstrates four quantitative developmental-biology tasks:
#
# 1. Estimate early exponential growth rate and doubling time.
# 2. Simulate constrained logistic growth.
# 3. Model branching differentiation into two lineages.
# 4. Assign developmental fates from a morphogen-gradient threshold.
#
# These examples can be adapted for embryology, plant development,
# stem cell biology, organoid systems, regenerative biology,
# developmental ecology, tissue engineering, and computational biology.
library(dplyr)
library(tidyr)
library(tibble)
# ------------------------------------------------------------
# 1. Developmental growth rate and doubling time
# ------------------------------------------------------------
growth_df <- tibble(
time_h = c(0, 6, 12, 18, 24, 30, 36),
cells = c(1.0e4, 1.4e4, 2.0e4, 2.8e4, 4.0e4, 5.1e4, 5.8e4)
)
# Fit the early approximately exponential phase.
fit_exp <- lm(log(cells) ~ time_h, data = growth_df %>% slice(1:5))
r_est <- coef(fit_exp)[["time_h"]]
N0_est <- exp(coef(fit_exp)[["(Intercept)"]])
doubling_time_h <- log(2) / r_est
growth_summary <- tibble(
r_est = r_est,
N0_est = N0_est,
doubling_time_h = doubling_time_h
)
growth_df <- growth_df %>%
mutate(
exp_pred = exp(predict(fit_exp, newdata = growth_df)),
residual = cells - exp_pred
)
# ------------------------------------------------------------
# 2. Logistic growth under developmental constraint
# ------------------------------------------------------------
simulate_logistic <- function(times, N0 = 1e4, r = 0.07, K = 6.2e4) {
N <- numeric(length(times))
N[1] <- N0
for (i in 2:length(times)) {
dt <- times[i] - times[i - 1]
dN <- r * N[i - 1] * (1 - N[i - 1] / K)
N[i] <- N[i - 1] + dN * dt
}
tibble(time_h = times, logistic_N = N)
}
logistic_df <- simulate_logistic(seq(0, 40, by = 0.25))
# ------------------------------------------------------------
# 3. Branching differentiation into two lineages
# ------------------------------------------------------------
simulate_lineage_split <- function(times, P0 = 1.0, k1 = 0.18, k2 = 0.10) {
P <- numeric(length(times))
D1 <- numeric(length(times))
D2 <- numeric(length(times))
P[1] <- P0
D1[1] <- 0
D2[1] <- 0
for (i in 2:length(times)) {
dt <- times[i] - times[i - 1]
dP <- -(k1 + k2) * P[i - 1]
dD1 <- k1 * P[i - 1]
dD2 <- k2 * P[i - 1]
P[i] <- max(P[i - 1] + dP * dt, 0)
D1[i] <- D1[i - 1] + dD1 * dt
D2[i] <- D2[i - 1] + dD2 * dt
}
tibble(
time = times,
progenitor = P,
lineage_1 = D1,
lineage_2 = D2
)
}
lineage_df <- simulate_lineage_split(seq(0, 25, by = 0.1))
lineage_long <- lineage_df %>%
pivot_longer(
cols = -time,
names_to = "state",
values_to = "fraction"
)
# ------------------------------------------------------------
# 4. Morphogen gradient and threshold-based fate assignment
# ------------------------------------------------------------
x <- seq(0, 1, length.out = 400)
morphogen <- exp(-5 * x)
fate_df <- tibble(
position = x,
morphogen = morphogen
) %>%
mutate(
fate = case_when(
morphogen > 0.60 ~ "fate_A",
morphogen > 0.25 ~ "fate_B",
TRUE ~ "fate_C"
)
)
fate_summary <- fate_df %>%
count(fate)
print(round(growth_summary, 4))
print(round(growth_df, 2))
print(logistic_df %>% slice_head(n = 12) %>% mutate(logistic_N = round(logistic_N, 2)))
print(logistic_df %>% slice_tail(n = 12) %>% mutate(logistic_N = round(logistic_N, 2)))
print(lineage_df %>% slice_head(n = 12) %>% mutate(across(where(is.numeric), round, 4)))
print(lineage_df %>% slice_tail(n = 12) %>% mutate(across(where(is.numeric), round, 4)))
print(lineage_long %>% slice_head(n = 12) %>% mutate(fraction = round(fraction, 4)))
print(fate_df %>% slice_head(n = 12) %>% mutate(morphogen = round(morphogen, 4)))
print(fate_summary)This R workflow is useful because developmental growth is rarely purely exponential throughout an entire process. A scientist-facing treatment should show early proliferation, later constraint, branching lineage allocation, and positional-information logic together.
Python workflow: branching differentiation, reaction-diffusion, stage growth, and state transitions
Python is useful for developmental biology because it supports numerical simulation, matrix operations, state-transition models, reaction-diffusion systems, pipeline design, and reproducible computation. The following workflow simulates branching differentiation, one-dimensional reaction-diffusion patterning, stage-dependent logistic growth, and Markov-style developmental state transitions.
"""
Development, Differentiation, and the Making of Organisms Workflow
This workflow demonstrates four quantitative developmental-biology tasks:
1. Simulate branching differentiation from a progenitor pool.
2. Simulate a one-dimensional reaction-diffusion pattern.
3. Simulate logistic growth with a developmental stage shift.
4. Model developmental state transitions with a Markov-style matrix.
The examples are compact, but the same structures can be extended to
embryology, plant development, stem cell biology, organoid systems,
regenerative biology, developmental ecology, tissue engineering,
and computational developmental biology.
"""
from __future__ import annotations
import numpy as np
import pandas as pd
def simulate_branching_differentiation(
t_end: float = 30.0,
dt: float = 0.1,
progenitor0: float = 1000.0,
k1: float = 0.14,
k2: float = 0.09,
) -> pd.DataFrame:
"""
Simulate progenitor decline into two differentiated lineages.
"""
times = np.arange(0.0, t_end + dt, dt)
progenitor = np.zeros_like(times)
lineage_1 = np.zeros_like(times)
lineage_2 = np.zeros_like(times)
progenitor[0] = progenitor0
for i in range(1, len(times)):
dP = -(k1 + k2) * progenitor[i - 1]
dD1 = k1 * progenitor[i - 1]
dD2 = k2 * progenitor[i - 1]
progenitor[i] = max(progenitor[i - 1] + dP * dt, 0.0)
lineage_1[i] = lineage_1[i - 1] + dD1 * dt
lineage_2[i] = lineage_2[i - 1] + dD2 * dt
return pd.DataFrame(
{
"time": times,
"progenitor": progenitor,
"lineage_1": lineage_1,
"lineage_2": lineage_2,
}
)
def simulate_reaction_diffusion(
n: int = 120,
steps: int = 1500,
Du: float = 0.001,
Dv: float = 0.01,
a: float = 0.04,
b: float = 0.065,
dt: float = 1.0,
seed: int = 42,
) -> pd.DataFrame:
"""
Simulate a simple one-dimensional activator-inhibitor pattern.
"""
rng = np.random.default_rng(seed)
u = np.ones(n) + 0.01 * rng.normal(size=n)
v = np.zeros(n) + 0.01 * rng.normal(size=n)
def laplacian(arr: np.ndarray) -> np.ndarray:
left = np.roll(arr, 1)
right = np.roll(arr, -1)
return left - 2.0 * arr + right
for _ in range(steps):
Lu = laplacian(u)
Lv = laplacian(v)
du = Du * Lu + a - u + u * u * v
dv = Dv * Lv + b - u * u * v
u += du * dt
v += dv * dt
return pd.DataFrame(
{
"position": np.arange(n),
"u": u,
"v": v,
}
)
def stage_dependent_growth(
t_end: float = 40.0,
dt: float = 0.1,
N0: float = 1.0e4,
K: float = 8.0e4,
r_early: float = 0.08,
r_late: float = 0.03,
switch_time: float = 18.0,
) -> pd.DataFrame:
"""
Simulate logistic growth with a developmental stage shift.
"""
times = np.arange(0.0, t_end + dt, dt)
cells = np.zeros_like(times)
cells[0] = N0
for i in range(1, len(times)):
r = r_early if times[i - 1] < switch_time else r_late
dN = r * cells[i - 1] * (1.0 - cells[i - 1] / K)
cells[i] = cells[i - 1] + dN * dt
return pd.DataFrame(
{
"time": times,
"cells": cells,
}
)
def developmental_state_transition() -> pd.DataFrame:
"""
Simulate a Markov-style developmental state transition matrix.
"""
states = ["progenitor", "transitional", "lineage_A", "lineage_B"]
transition_matrix = np.array(
[
[0.76, 0.20, 0.03, 0.01],
[0.04, 0.72, 0.16, 0.08],
[0.00, 0.03, 0.94, 0.03],
[0.00, 0.02, 0.04, 0.94],
]
)
state_vector = np.array([0.90, 0.08, 0.01, 0.01])
trajectory = [state_vector.copy()]
for _ in range(20):
state_vector = state_vector @ transition_matrix
trajectory.append(state_vector.copy())
trajectory_df = pd.DataFrame(trajectory, columns=states)
trajectory_df["step"] = range(len(trajectory_df))
return trajectory_df
def main() -> None:
"""
Run compact developmental-biology workflows.
"""
branching_df = simulate_branching_differentiation()
pattern_df = simulate_reaction_diffusion()
growth_df = stage_dependent_growth()
state_df = developmental_state_transition()
print("Branching differentiation:")
print(branching_df.head(15).round(4).to_string(index=False))
print(branching_df.tail(15).round(4).to_string(index=False))
print("\nReaction-diffusion pattern summary:")
print(pattern_df.head(20).round(4).to_string(index=False))
print(pattern_df.describe().round(4).to_string())
print("\nStage-dependent growth:")
print(growth_df.head(20).round(2).to_string(index=False))
print(growth_df.tail(20).round(2).to_string(index=False))
print("\nDevelopmental state-transition trajectory:")
print(state_df.round(4).to_string(index=False))
if __name__ == "__main__":
main()This Python workflow is useful because development often requires dynamic modeling across cell states, spatial fields, growth regimes, and transition probabilities. It provides a practical scaffold for thinking about developmental trajectories rather than treating development as a static sequence of labels.
GitHub repository
The article body includes compact R and Python examples so the biological and scientific argument remains readable. The full repository expands those examples into a broader computational developmental-biology workflow, including exponential growth fitting, doubling-time estimation, logistic growth, stage-dependent growth, branching differentiation, morphogen-gradient thresholding, one-dimensional reaction-diffusion patterning, developmental state-transition modeling, organoid-style developmental condition scoring, SQL provenance structures, reproducible data files, validation notes, and full-stack scientific-computing examples across Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, and notebooks.
Complete Code Repository
The full code distribution for this article, including selected article examples, expanded computational workflows, reproducible data structures, provenance documentation, validation notes, and full-stack scientific-computing scaffolding, is available on GitHub.
Limits, complexity, and modern developmental thinking
Developmental biology is foundational, but development is not simple. The same genome can support many developmental outcomes depending on regulation, timing, environment, signaling, mechanics, cell history, and interaction. Similar structures can arise through different routes, and similar perturbations can have very different effects depending on developmental stage. A developmental process characterized cleanly in one model organism may not map perfectly onto another.
This is why modern developmental thinking increasingly emphasizes integration across genetics, molecular biology, cell biology, signaling, mechanics, ecology, evolution, and computation. Development is strongest as an explanatory field when it remains multi-scale. It cannot be reduced to genes alone, cells alone, tissue mechanics alone, environmental exposure alone, or mathematical models alone.
Models are useful because they clarify assumptions, expose mechanisms, and make comparison possible. But a growth curve is not a whole embryo, a morphogen gradient is not the entire patterning system, and a reaction-diffusion simulation is not direct proof of a specific developmental mechanism. Quantitative tools are strongest when they support biological interpretation rather than replacing it.
In that sense, development provides one of the clearest models of modern biology itself: dynamic, historically informed, mechanistically rich, environmentally responsive, quantitatively analyzable, and irreducible to a single level of explanation. Its strength lies in showing how life builds itself through coordinated process rather than presupposed final form.
This caution matters especially in high-throughput developmental biology. A cell-state trajectory may look precise but still depend on sampling time, normalization, clustering choices, marker interpretation, spatial context, and biological validation. Developmental evidence is strongest when computational inference, experimental perturbation, lineage tracing, imaging, and functional observation converge.
Why this matters for scientific work
For working scientists, development matters because many biological problems are misread when organisms are treated as if their current form were simply given rather than produced. A conservation problem may actually be a recruitment problem. A physiological problem may begin in tissue organization laid down developmentally. A restoration problem may depend on germination and juvenile establishment windows. A disease problem may reflect failed differentiation or abnormal developmental timing rather than only adult malfunction.
This means developmental biology should often be treated as explanatory infrastructure rather than as a specialized corner of embryology. Ecologists need it because survival often depends on developmental stage. Evolutionary biologists need it because variation is filtered through developmental possibility. Plant scientists need it because growth and form are developmentally organized throughout the life cycle. Marine and freshwater biologists need it because early life stages can determine whole population trajectories. Biomedical scientists need it because tissues, pathologies, and regenerative capacities all depend on developmental logic. Computational biologists need it because development is one of biology’s most dynamic and structured time-series problems.
The scientific importance of development lies partly in this breadth. It is one of the principal ways biology explains how living systems become what they are.
Development is also practically actionable. Growth rates can be measured. Differentiation trajectories can be modeled. Stem-cell protocols can be optimized. Morphogen responses can be tested. Developmental toxicity can be screened. Recruitment bottlenecks can be studied. Regeneration can be compared. These tools connect developmental theory to medicine, conservation, agriculture, biotechnology, ecology, and systems biology.
Conclusion
Development, differentiation, and the making of organisms show that life is not merely inherited as static form. It is produced through regulated processes of specification, differentiation, morphogenesis, patterning, growth, and life-cycle coordination. Organisms emerge because cells acquire distinct identities, communicate across space and time, and organize themselves into functional structures.
To understand development is therefore to understand one of the deepest conditions of biological form. Development links heredity to anatomy, regulation to physiology, cellular process to organismal identity, and environmental condition to embodied outcome. That is why developmental biology remains central not only to embryology and cell differentiation, but also to ecology, evolution, plant science, marine and freshwater biology, disease ecology, medicine, biotechnology, and sustainability-adjacent biology more broadly.
Development is thus more than a branch of biology. It is one of the principal ways biology explains how living systems become what they are. Modern quantitative and computational workflows deepen that understanding by making growth, differentiation, patterning, state transitions, and developmental provenance more transparent, reproducible, and scientifically interpretable.
Related articles
- Biology
- Cell Theory and the Basic Unit of Life
- DNA, RNA, and the Molecular Logic of Life
- Molecular Biology and the Flow of Genetic Information
- Cell Signaling, Communication, and Biological Coordination
- Genes, Inheritance, and the Principles of Heredity
- Epigenetics, Regulation, and Gene Expression
- Physiology and the Regulation of Living Systems
- Animal Biology and the Organization of Complex Life
- Plant Biology and the Life of Primary Producers
- Evolution and the History of Life
- Systems Biology and the Logic of Biological Integration
Further reading
- Gilbert, S.F. (2000) Developmental Biology. 6th edn. Sunderland, MA: Sinauer Associates. NCBI Bookshelf edition available at: https://www.ncbi.nlm.nih.gov/books/NBK9983/
- Gilbert, S.F. (2000) ‘An introduction to early developmental processes’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9992/
- Gilbert, S.F. (2000) ‘The developmental mechanics of cell specification’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9968/
- Gilbert, S.F. (2000) ‘Morphogenesis and cell adhesion’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10021/
- Gilbert, S.F. (2000) ‘Multicellularity: The evolution of differentiation’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10031/
- National Human Genome Research Institute (n.d.) Stem Cell. Available at: https://www.genome.gov/genetics-glossary/Stem-Cell
- National Research Council (2000) ‘Development’, in Opportunities in Biology. Washington, DC: National Academies Press. Available at: https://www.ncbi.nlm.nih.gov/books/NBK217800/
- Turing, A.M. (1952) ‘The chemical basis of morphogenesis’, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 237(641), pp. 37–72. Available at: https://doi.org/10.1098/rstb.1952.0012
- Vargesson, N. (2020) ‘Positional information: A concept underpinning our understanding of patterning in development’, Developmental Dynamics, 249(3), pp. 287–297. Available at: https://pubmed.ncbi.nlm.nih.gov/31566855/
- Wolpert, L. (2016) ‘Positional information and pattern formation’, Current Topics in Developmental Biology, 117, pp. 597–608. Available at: https://pubmed.ncbi.nlm.nih.gov/26970003/
References
- Gilbert, S.F. (2000) Developmental Biology. 6th edn. Sunderland, MA: Sinauer Associates. NCBI Bookshelf edition available at: https://www.ncbi.nlm.nih.gov/books/NBK9983/
- Gilbert, S.F. (2000) ‘An introduction to early developmental processes’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9992/
- Gilbert, S.F. (2000) ‘The developmental mechanics of cell specification’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9968/
- Gilbert, S.F. (2000) ‘Morphogenesis and cell adhesion’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10021/
- Gilbert, S.F. (2000) ‘Multicellularity: The evolution of differentiation’, in Developmental Biology. 6th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10031/
- National Human Genome Research Institute (n.d.) Stem Cell. Available at: https://www.genome.gov/genetics-glossary/Stem-Cell
- National Research Council (2000) ‘Development’, in Opportunities in Biology. Washington, DC: National Academies Press. Available at: https://www.ncbi.nlm.nih.gov/books/NBK217800/
- Turing, A.M. (1952) ‘The chemical basis of morphogenesis’, Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 237(641), pp. 37–72. Available at: https://doi.org/10.1098/rstb.1952.0012
- Vargesson, N. (2020) ‘Positional information: A concept underpinning our understanding of patterning in development’, Developmental Dynamics, 249(3), pp. 287–297. Available at: https://pubmed.ncbi.nlm.nih.gov/31566855/
- Wolpert, L. (2016) ‘Positional information and pattern formation’, Current Topics in Developmental Biology, 117, pp. 597–608. Available at: https://pubmed.ncbi.nlm.nih.gov/26970003/