
Last Updated May 28, 2026
Immunology and biological defense examine how living systems detect danger, distinguish self from non-self or altered self, coordinate protective responses against infection and damage, remember prior exposure, and regulate the fine balance between defense, tolerance, inflammation, repair, and injury. Immunology is central to biology because life persists not only through metabolism, development, reproduction, and homeostasis, but also through the capacity to resist invasion, contain damage, regulate microbial relationships, and preserve internal integrity under continual environmental challenge. Biological defense is not merely an emergency reaction. It is one of the permanent conditions of living organization.
Immunology is therefore one of biology’s great integrative sciences. It connects molecular recognition to cellular activation, tissue barriers to organismal physiology, inflammation to repair, memory to future protection, microbiology to host ecology, vaccination to population health, and disease dynamics to evolutionary pressure. The immune system is not a simple weapon system aimed outward at foreign material. It is a distributed regulatory system that must decide when to attack, when to tolerate, when to remember, when to repair, and when to stop.
Main Library
Publications
Article Map
Biology
Related Topic
Physiology
Related Topic
Microbiology
Related Topic
Ecology
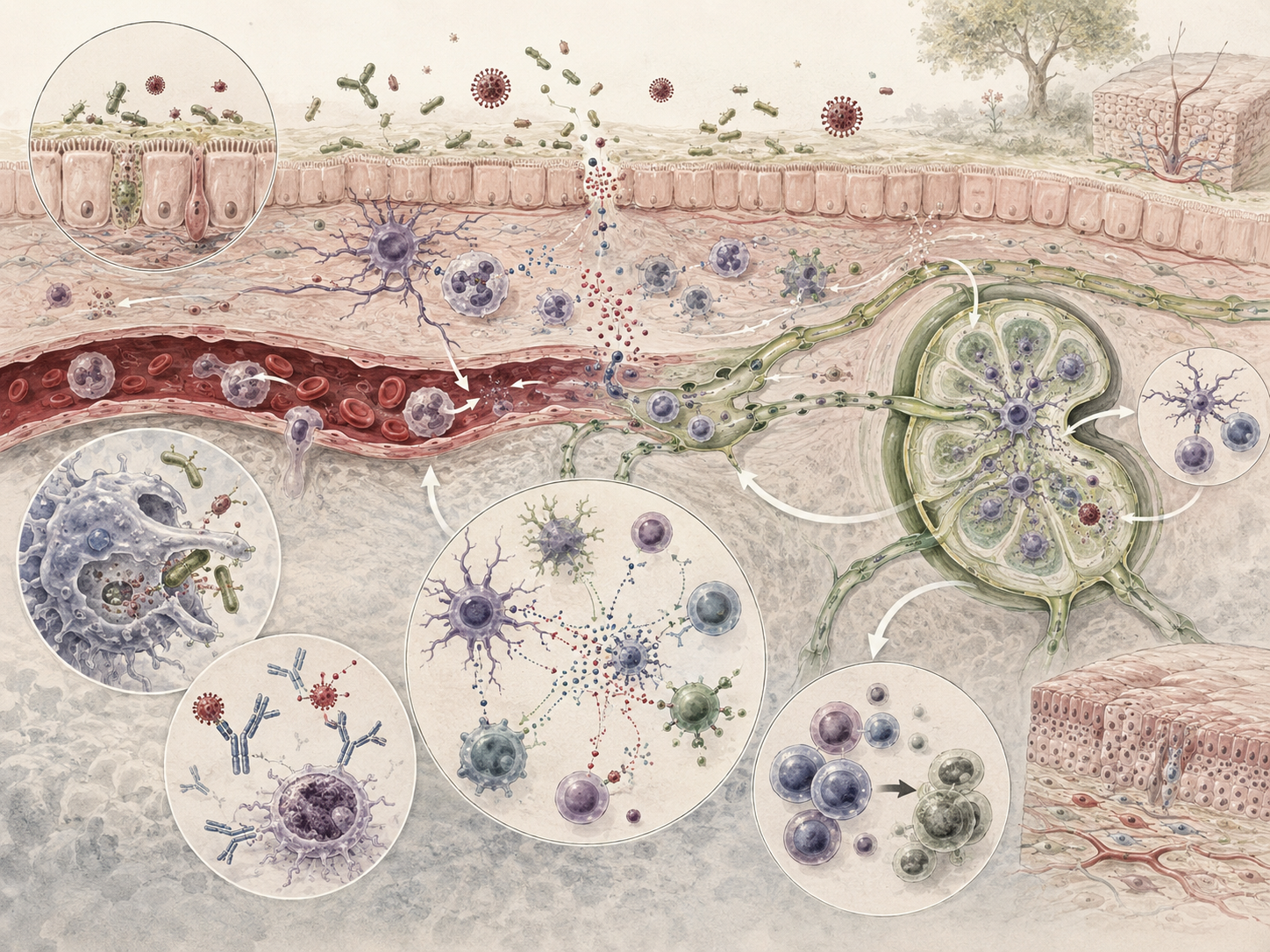
This article develops immunology and biological defense as a scale-spanning framework for understanding living protection. It examines innate immunity, adaptive immunity, pattern recognition, barriers, phagocytes, lymphocytes, antibodies, complement, cytokines, chemokines, inflammation, tolerance, memory, vaccination, host–pathogen interaction, immune dysregulation, comparative defense across animals, plants, and microbes, disease ecology, conservation immunology, systems immunology, genomics, and computational modeling.
The article is written for immunologists, microbiologists, ecologists, marine biologists, freshwater scientists, medical and environmental-health readers, computational biology readers, biotech readers, biodiversity experts, conservation planners, plant scientists, animal biologists, systems biologists, and research biologists who need a rigorous account of how defense systems operate from molecules to tissues, organisms, populations, and ecological systems.
The article also extends immunology into quantitative and computational biology through host–pathogen dynamics, inflammatory feedback models, immune activation and resolution systems, tissue-damage trade-offs, threshold screening, scenario analysis, R workflows, Python workflows, SQL provenance structures, and a linked full-stack GitHub repository containing Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, notebooks, data files, and reproducibility documentation.
What immunology studies
Immunology is the study of biological defense: how organisms recognize and respond to pathogens, toxins, tissue damage, abnormal cells, parasites, microbial disruption, and other threats to internal integrity. The immune system consists of barriers, cells, tissues, soluble molecules, receptors, signaling networks, and memory mechanisms that provide innate and acquired protection. Proper immune function requires not only attack, but discrimination, timing, regulation, proportionality, and context-sensitive judgment.
This matters because life must remain open enough to exchange matter and energy with the environment while also remaining sufficiently defended against invasion and internal disorder. Organisms breathe, eat, absorb, reproduce, interact, and maintain resident microbes, but each of those openings creates exposure. Immunology studies that tension. It asks how organisms protect themselves without destroying their own tissues, tolerate what must be tolerated, and respond strongly enough to survive.
Immunology also studies failure. Infection, sepsis, chronic inflammation, allergy, autoimmunity, immunodeficiency, immune evasion, cancer immune escape, transplant rejection, vaccine failure, and immune exhaustion all reveal that defense is a regulated system rather than a simple on-off mechanism. Too little defense permits invasion. Too much or misdirected defense damages the host.
For research biologists, immunology belongs near the center of biology because it links physiology, ecology, microbiology, development, evolution, medicine, and environmental stress into one dynamic science of regulated defense.
Biological defense and the problem of self, nonself, and damage
One of immunology’s most important themes is discrimination: how organisms distinguish self from non-self, harmless from dangerous, resident from invasive, tolerable from destructive, and damaged from healthy. Proper immune function requires distinction between the body’s own material and material originating outside it, but it also requires distinguishing among harmless antigens, commensal microbes, damaged tissues, abnormal cells, reproductive tissues, food molecules, and true infectious threat.
This matters because biological defense cannot simply attack everything unfamiliar. Organisms contain beneficial microbes, environmental exposures, dietary molecules, developing structures, and damaged but recoverable cells that must not be treated identically to pathogens. Immune systems therefore do more than destroy invaders. They classify biological situations.
Self/non-self recognition remains important, but modern immunology also recognizes that context, damage, location, molecular pattern, tissue state, inflammatory signaling, and regulatory feedback all influence immune interpretation. A molecule may be harmless in one context and dangerous in another. A microbe may be tolerated in one tissue and dangerous in another. A strong response may be protective early and damaging later.
Immunology is strongest when it is understood not only as warfare, but as regulated judgment under uncertainty. Defense depends on recognition, context, proportionality, memory, tolerance, repair, and the continual management of false positives and false negatives.
Innate immunity: the first line of defense
Innate immunity is the rapid, broadly acting arm of biological defense. It is the body’s first line of defense, responding quickly to microbes, toxins, foreign substances, tissue injury, and danger-associated signals. Barriers, phagocytes, complement, inflammatory mediators, antimicrobial peptides, natural killer cells, epithelial defenses, and pattern-recognition systems all belong to this level.
This matters because organisms often cannot wait for highly tailored responses before acting. Pattern-recognition receptors such as Toll-like receptors detect common signatures of microbial infection and initiate early protective cascades. Macrophages, neutrophils, dendritic cells, epithelial cells, and complement pathways help contain threat immediately, buying time and often resolving danger before highly specific responses fully emerge.
Innate immunity is often described as nonspecific, but that word can be misleading if it implies crude or simple defense. Innate systems can distinguish broad molecular patterns, tissue contexts, damage signals, microbial categories, and inflammatory states. They may not generate the same receptor diversity and clonal memory characteristic of adaptive immunity, but they are not biologically naive.
For research biologists, innate immunity reveals that defense begins with ancient, generalizable recognition systems. It is the fast architecture of survival, and it connects directly to evolution, microbiology, physiology, and ecological exposure.
Adaptive immunity: specificity, memory, and lymphocyte response
Adaptive immunity is the more specific and memory-forming arm of vertebrate defense. Adaptive responses are slower to initiate than many innate responses, but they provide receptor specificity, clonal expansion, antibody production, T-cell-mediated defense, and durable immunological memory. B cells and T cells enable the body to distinguish and react to a vast range of foreign substances through receptor-based recognition, differentiation, and effector function.
This matters because many pathogens cannot be controlled adequately by broad innate defenses alone. Adaptive immunity allows hosts to generate more selective responses, including antibody production, cytotoxic T-cell activity, helper T-cell coordination, affinity maturation, plasma-cell production, and memory-cell persistence. Defense therefore becomes historical: the host is changed by encounter.
Adaptive immunity is also a biological information system. Lymphocyte repertoires represent enormous recognition potential. Clonal expansion amplifies rare cells with useful specificity. Memory preserves traces of prior exposure. Vaccination uses this logic deliberately by training immune memory without requiring the full burden of natural infection.
For research biologists, adaptive immunity is one of the clearest examples of biological information storage through experience. The immune system does not merely react. It remembers, refines, and prepares.
Innate and adaptive immunity as an integrated system
Innate and adaptive immunity are often taught separately, but in living systems they are deeply integrated. Most infectious agents induce inflammatory responses by activating innate immunity, and these innate responses help initiate adaptive immunity through antigen presentation, cytokine signaling, costimulation, lymphocyte recruitment, and lymphoid-tissue organization. Dendritic cells, macrophages, cytokine networks, complement fragments, and lymphoid tissues help link these levels of response.
This matters because biological defense is not a sequence of isolated modules. Rapid recognition, inflammatory recruitment, antigen processing, antigen presentation, lymphocyte activation, effector function, resolution, and immune memory are parts of one coordinated system. Timing, location, and signaling architecture matter as much as the individual components themselves.
The integrated character of immunity is especially important in infection, vaccination, cancer immunology, autoimmunity, allergy, chronic inflammation, and immunodeficiency. A defect in innate sensing can impair adaptive activation. A dysregulated adaptive response can prolong inflammation. A failure of regulation can turn protective defense into pathology.
For research biologists, this makes immunology a systems science. Defense works because different temporal and functional layers are coupled rather than because one layer alone is sufficient.
Barriers, cells, and molecules of defense
Biological defense includes physical barriers, specialized cells, and soluble molecules. Skin and mucosal surfaces form protective boundaries. Epithelial cells participate actively in defense through secretions, receptor signaling, and barrier regulation. Phagocytes engulf invaders and debris. Natural killer cells target abnormal or infected cells. Lymphocytes drive specific recognition and memory. Antibodies, complement proteins, cytokines, chemokines, interferons, antimicrobial peptides, acute-phase proteins, and tissue mediators shape the chemistry of defense.
This matters because immune defense is not reducible to one cell type, one organ, or one molecular pathway. It is distributed across blood, lymph, tissues, epithelia, bone marrow, thymus, spleen, lymph nodes, mucosal sites, microbiomes, and signaling molecules. The immune system is best understood as a network of surveillance, communication, recruitment, regulation, and response extending across the body.
Barriers are especially important because they show that immunity is not only a reaction after invasion. Much biological defense is preventive. Skin, mucus, cilia, stomach acid, antimicrobial secretions, tissue architecture, microbiome competition, and epithelial turnover all contribute to protection before immune escalation becomes necessary.
For research biologists, this is one reason immunology belongs closely with physiology. Defense is one of the great regulatory functions of life, and it is distributed rather than localized in any one place.
Pattern recognition, danger, and immune classification
Immune systems depend on recognition. Pattern-recognition receptors detect molecular signatures associated with microbes, damaged cells, or altered tissue states. These include pathogen-associated molecular patterns, damage-associated molecular patterns, and other contextual signals that help immune systems classify biological situations. Toll-like receptors are among the most studied examples, but they are part of a broader landscape of innate sensors.
This matters because immune response begins before the organism has a full narrative of what is happening. The immune system detects molecular evidence and acts under uncertainty. A microbial signature, cellular stress signal, foreign nucleic acid, damaged membrane, altered protein, or inflammatory mediator can initiate cascades that recruit cells and shape later adaptive immunity.
Pattern recognition also helps explain why innate immunity is not merely primitive. It is a sophisticated classification system built from evolutionary experience. It does not need to identify every individual pathogen species in order to respond meaningfully. It recognizes recurring molecular patterns that have defensive significance.
For research biologists, pattern recognition is one of the strongest bridges between molecular biology, evolution, infection, inflammation, and systems-level immune control.
Inflammation, tolerance, and the double edge of protection
Inflammation is one of the immune system’s most important responses because it helps recruit cells, alter vascular behavior, mobilize mediators, localize defense, and begin repair at sites of infection or damage. Acute inflammation can be protective when it contains threat and supports healing. Yet inflammation is also dangerous when excessive, misdirected, unresolved, or chronic. A response too weak permits infection; a response too strong damages host tissues.
This matters because immune protection is double-edged. Tolerance is not passive failure but an active necessity: organisms must tolerate food antigens, commensal microbes, self tissues, reproductive tissues, harmless environmental inputs, and many low-risk exposures while remaining ready to respond when danger appears. Regulation is therefore as central to immunology as activation.
Immune regulation includes anti-inflammatory cytokines, regulatory T cells, checkpoint pathways, tissue repair programs, tolerance mechanisms, clearance of apoptotic cells, feedback inhibition, and the spatial confinement of inflammatory responses. These processes are not secondary. They are essential to survival.
For research biologists, this is one of the deepest lessons in the field: defense is not simply attack. It is calibrated response under the risk of self-inflicted injury.
Immune memory, vaccination, and long-term protection
One of adaptive immunity’s most important features is memory: prior exposure can lead to more rapid, specific, and effective later responses. Acquired immunity and vaccination both rely on this principle. The host is altered by encounter, and later immune responses can be faster, stronger, or more precisely targeted.
This matters because immune memory makes defense cumulative rather than purely momentary. Vaccination takes advantage of this by inducing protective memory without requiring the full burden of natural disease. Memory B cells, memory T cells, long-lived plasma cells, and antibody persistence all contribute to protection in different ways.
Immune memory is also a reminder that biology is historical. The immune system is shaped by previous infection, vaccination, microbiome exposure, inflammation, age, nutrition, stress, and environmental conditions. No immune system exists outside its history.
For research biologists, immune memory is especially important because it links molecular recognition, cellular persistence, developmental fate, population-level public health, and evolutionary pressure. It is one of biology’s clearest examples of adaptive protection through prior information.
Immunology in animals, plants, and microbes
Although immunology is often taught through vertebrate examples, biological defense is broader than vertebrate adaptive immunity alone. All multicellular organisms must defend against infection and damage, and plants and microbes also possess powerful defense systems. Plants detect pathogen-associated signals and activate layered immune responses. Invertebrates rely heavily on innate defenses, cellular responses, antimicrobial molecules, melanization, and barrier systems. Microbes possess defense systems against phages, mobile genetic elements, and other threats.
This matters because biological defense is a general problem of life, not a narrow specialty of human medicine. Different lineages solve it differently, but the challenge of detecting, resisting, regulating, and surviving threat is widespread. Defense is one of the main ways living systems preserve continuity in microbial worlds.
Plant immunity is especially important because it shows that defense can be highly sophisticated without vertebrate-style antibodies or lymphocytes. Microbial defense is equally important because it shows that even bacteria exist in worlds of invasion, parasitism, and molecular conflict.
For research biologists, comparative immunology is valuable because it shows both the diversity and recurrence of defensive design across life. The details differ, but the problem of biological defense recurs.
Host–pathogen coevolution and disease ecology
Immune systems evolve in relation to pathogens, and pathogens evolve in relation to host defense. Hosts refine recognition, containment, memory, tissue protection, and tolerance; pathogens evolve evasion, concealment, rapid replication, immune modulation, antigenic change, latency, host manipulation, or exploitation of host signaling. Immunology is therefore inherently coevolutionary.
This matters because disease emergence, virulence, transmission, reservoir dynamics, spillover, and epidemic behavior all depend on the interplay among host defenses, pathogen capacities, environment, and population structure. Immune systems are shaped by microbial worlds, not merely by internal design. Ecological context changes which threats are encountered, how they are transmitted, and which defensive strategies are favored.
Disease ecology also shows that immunity is not only an individual property. Population density, social behavior, migration, climate, land use, biodiversity, vector ecology, stress, nutrition, and habitat disturbance can all alter exposure and immune outcomes. Biological defense scales from molecules to ecosystems.
For research biologists, this makes immunology inseparable from ecology and evolution. Defense is never static because pathogens are not static, and environments continually alter the terms of encounter.
Marine, freshwater, soil, and terrestrial defense systems
Defense systems operate under very different ecological pressures across marine, freshwater, soil, and terrestrial environments. Aquatic organisms face distinct pathogen loads, salinity and temperature stresses, osmotic constraints, dissolved oxygen variation, transmission pathways, and microbial exposures. Soil organisms live amid dense microbial communities and chemically complex substrates. Terrestrial organisms encounter different barrier challenges, vector exposures, dehydration risks, airborne pathogens, and environmental fluctuations.
This matters because immunity is always situated. A fish, amphibian, coral, plant root, insect, bird, mammal, fungus, or soil microbe does not defend itself in the abstract. Defense is shaped by habitat, life history, symbiosis, barrier structure, microbiome composition, temperature, stress physiology, and exposure regime.
Marine and freshwater immunology are especially relevant under climate change, pollution, hypoxia, acidification, harmful algal blooms, and habitat alteration. Soil and plant defense matter for agroecology, forestry, restoration ecology, and food systems. Terrestrial wildlife immunology matters for conservation, zoonotic disease, and environmental health.
For research biologists, ecological situatedness matters deeply. Climate stress, pollution, habitat disruption, and changing microbial communities can all alter disease risk by altering biological defense.
Immunology, conservation, and systems-oriented biology
Immunology matters for conservation because populations under environmental change often face immune stress before they face extinction. Heat, malnutrition, pollutants, endocrine disruption, salinity shifts, drought, fragmentation, crowding, migration disruption, and altered microbial communities can all change susceptibility to infection or inflammatory damage. Immune function is therefore part of population resilience.
This matters because conservation problems are not only about abundance or habitat cover. They are also about whether organisms and populations can maintain functional defense under changing conditions. Disease outbreaks in wildlife, coral stress, amphibian decline, crop vulnerability, pollinator pathogens, livestock-wildlife interfaces, and soil community disruption all have immunological dimensions.
Conservation immunology asks how immune function varies across individuals, populations, species, environments, and stress regimes. It connects physiology to population viability and disease ecology. A population may remain numerically present while becoming immunologically compromised, more susceptible to disease, or less able to recover from environmental stress.
For research biologists, immunology provides one of the strongest bridges between organismal biology and resilience under environmental change. It clarifies how stress becomes susceptibility and how susceptibility can scale into ecological consequence.
Systems immunology, genomics, and computational relevance
Modern immunology increasingly depends on genomics, transcriptomics, proteomics, metabolomics, single-cell profiling, cell atlases, signaling-network analysis, mathematical modeling, and computational inference. Immune responses are now understood not only through individual cell types or molecules, but as networked and time-dependent systems involving recognition, recruitment, differentiation, metabolism, spatial organization, memory, and tissue context.
This matters because immune states emerge through interacting layers of recognition, signaling, recruitment, differentiation, regulation, metabolism, repair, and history. Systems immunology helps make that complexity legible by treating defense as a dynamic system rather than a list of components.
Computational approaches can help model infection trajectories, immune-cell expansion, cytokine dynamics, tissue damage, vaccine response, immune exhaustion, clonal repertoires, antigen recognition, and host-pathogen interaction. But those models must remain connected to biological interpretation. An immune score is not the immune system itself. A differential equation is not a tissue. A cell atlas is not the whole organism.
For research biologists, immunology is one of the most integrative areas of modern biology. It sits naturally at the intersection of classical host defense, molecular biology, ecology, medicine, and contemporary computational life science.
Quantitative immunology: mathematics, R, and Python
Immunology is deeply quantitative because defense depends on rates of pathogen growth, immune-cell expansion, signaling thresholds, cytokine decay, tissue damage, repair, regulation, memory formation, and feedback control. A simple conceptual model for pathogen burden under immune control can begin with:
\frac{dP}{dt}=rP-cIP
\]
Interpretation: \(P\) is pathogen load, \(r\) is pathogen growth rate, \(I\) is immune activity, and \(c\) is the clearance coefficient. This captures the broad idea that pathogen burden rises through replication and falls through immune-mediated control.
A second compact model for inflammatory regulation can be written as:
\frac{dX}{dt}=aS-bX
\]
Interpretation: \(X\) is inflammatory activity, \(S\) is stimulus intensity, \(a\) is activation strength, and \(b\) is the damping or resolution term. This is useful because immune responses must often both activate and resolve.
For research biologists, however, real immunology usually requires more structure. A minimal coupled host–pathogen–immune system can be written as:
\frac{dP}{dt}=rP-cIP
\]
Interpretation: Pathogen load increases through replication and decreases through immune-mediated clearance.
\frac{dI}{dt}=\alpha P-\delta I
\]
Interpretation: Immune activity increases in response to pathogen burden and declines through decay, regulation, or resolution.
\frac{dD}{dt}=\gamma I-\rho D
\]
Interpretation: Tissue damage or inflammatory burden increases with immune activity and declines through repair or resolution. This makes explicit one of immunology’s core truths: defense may reduce infection while also generating injury.
Worked example: net change in pathogen load
Suppose pathogen load is \(P=100\), pathogen growth rate is \(r=0.4\), immune activity is \(I=2\), and clearance coefficient is \(c=0.15\). Then:
\frac{dP}{dt}=0.4(100)-0.15(2)(100)=40-30=10
\]
Interpretation: Pathogen load is still rising under the simplified model, but more slowly than it would without immune clearance. The biological question then becomes whether immune activity can intensify rapidly enough to reverse that rise before damage becomes severe.
R and Python workflows
The following examples are compact article-level workflows. The full GitHub repository expands them into richer multi-language implementations with SQL provenance, validation notes, host–pathogen dynamics, immune activation and clearance, inflammatory damage, repair and resolution, threshold screening, immune-condition scoring, and reproducible computational immunology scaffolding.
R example: coupled host–pathogen–immune dynamics
# Coupled immunology model in R
#
# This compact workflow models:
# - pathogen load
# - immune activity
# - inflammatory or tissue-damage burden
# - repair or resolution
#
# Euler integration is used for article-scale clarity.
# This is a teaching scaffold, not a calibrated clinical or ecological model.
dt <- 0.05
time <- seq(0, 30, by = dt)
# Parameters
r <- 0.45 # pathogen growth
c <- 0.12 # immune clearance
alpha <- 0.08 # immune activation by pathogen
delta <- 0.18 # immune decay
gamma <- 0.06 # damage generated by immune activity
rho <- 0.10 # repair or resolution
# Initial states
P <- numeric(length(time))
I <- numeric(length(time))
D <- numeric(length(time))
P[1] <- 50
I[1] <- 2
D[1] <- 0
for (t in 2:length(time)) {
dP <- r * P[t - 1] - c * I[t - 1] * P[t - 1]
dI <- alpha * P[t - 1] - delta * I[t - 1]
dD <- gamma * I[t - 1] - rho * D[t - 1]
P[t] <- max(0, P[t - 1] + dP * dt)
I[t] <- max(0, I[t - 1] + dI * dt)
D[t] <- max(0, D[t - 1] + dD * dt)
}
results <- data.frame(
time = time,
pathogen_load = P,
immune_activity = I,
damage_burden = D
)
matplot(
time,
cbind(P, I, D),
type = "l",
lty = 1,
lwd = 2,
xlab = "Time",
ylab = "State value",
main = "Coupled Host-Pathogen-Immune Dynamics"
)
legend(
"topright",
legend = c("Pathogen load", "Immune activity", "Damage burden"),
lty = 1,
lwd = 2,
bty = "n"
)
print(tail(round(results, 3), 10))
diagnostics <- data.frame(
peak_pathogen = max(P),
peak_immune = max(I),
peak_damage = max(D),
final_pathogen = tail(P, 1),
final_damage = tail(D, 1)
)
print(round(diagnostics, 3))This R workflow is more useful than a single exponential pathogen curve because it models three coupled processes at once: infection, immune activation, and collateral damage. A research biologist could adapt it for acute infection, inflammatory disease, immune suppression, comparative host defense, conservation physiology, or ecophysiological stress scenarios.
Python example: comparative immune scenarios and threshold screening
import numpy as np
import pandas as pd
def simulate_scenario(
P0=50,
I0=2,
D0=0,
r=0.45,
c=0.12,
alpha=0.08,
delta=0.18,
gamma=0.06,
rho=0.10,
T=30,
dt=0.05
):
"""Simulate coupled pathogen, immune, and damage dynamics."""
time = np.arange(0, T + dt, dt)
P = np.zeros(len(time))
I = np.zeros(len(time))
D = np.zeros(len(time))
P[0], I[0], D[0] = P0, I0, D0
for t in range(1, len(time)):
dP = r * P[t - 1] - c * I[t - 1] * P[t - 1]
dI = alpha * P[t - 1] - delta * I[t - 1]
dD = gamma * I[t - 1] - rho * D[t - 1]
P[t] = max(0, P[t - 1] + dP * dt)
I[t] = max(0, I[t - 1] + dI * dt)
D[t] = max(0, D[t - 1] + dD * dt)
return {
"final_pathogen": P[-1],
"final_immune": I[-1],
"final_damage": D[-1],
"peak_pathogen": P.max(),
"peak_immune": I.max(),
"peak_damage": D.max()
}
scenarios = {
"weak_clearance": {"c": 0.08, "alpha": 0.06},
"moderate_clearance": {"c": 0.12, "alpha": 0.08},
"strong_clearance": {"c": 0.18, "alpha": 0.10},
"hyperinflammatory": {"c": 0.14, "alpha": 0.12, "gamma": 0.12},
"immune_suppressed": {"c": 0.06, "alpha": 0.04, "delta": 0.24}
}
rows = []
for name, params in scenarios.items():
result = simulate_scenario(**params)
result["scenario"] = name
rows.append(result)
df = pd.DataFrame(rows)
df["risk_class"] = np.where(
(df["peak_pathogen"] > 200) | (df["peak_damage"] > 20),
"high-risk",
np.where(
(df["peak_pathogen"] > 100) | (df["peak_damage"] > 10),
"stressed",
"controlled"
)
)
print(df.round(3))This Python workflow is more useful because it compares multiple immune-response regimes rather than presenting one fixed trajectory. It allows the reader to see how changes in clearance, activation, suppression, or collateral-damage parameters alter infection outcome and tissue burden. That makes it adaptable to disease ecology, conservation physiology, comparative immunology, host–pathogen modeling, and scenario-based teaching.
Python example: immune-condition score under ecological stress
import numpy as np
import pandas as pd
# Synthetic population or site-level immune-condition data.
# Values are scaled between 0 and 1 for article clarity.
immune_sites = pd.DataFrame({
"site": ["reef_A", "wetland_B", "forest_C", "farm_D", "river_E"],
"nutrition_status": [0.72, 0.64, 0.81, 0.58, 0.69],
"pathogen_pressure": [0.44, 0.61, 0.32, 0.70, 0.55],
"pollution_stress": [0.48, 0.35, 0.22, 0.66, 0.59],
"thermal_stress": [0.76, 0.41, 0.28, 0.52, 0.47],
"barrier_integrity": [0.62, 0.70, 0.84, 0.55, 0.60],
"inflammatory_burden": [0.58, 0.49, 0.31, 0.72, 0.64]
})
immune_sites["immune_condition_score"] = (
0.25 * immune_sites["nutrition_status"]
+ 0.25 * immune_sites["barrier_integrity"]
- 0.15 * immune_sites["pathogen_pressure"]
- 0.15 * immune_sites["pollution_stress"]
- 0.10 * immune_sites["thermal_stress"]
- 0.10 * immune_sites["inflammatory_burden"]
)
immune_sites["immune_risk_class"] = np.where(
immune_sites["immune_condition_score"] < 0.05,
"high-risk",
np.where(
immune_sites["immune_condition_score"] < 0.15,
"stressed",
"relatively-buffered"
)
)
print(immune_sites.round(3).to_string(index=False))This compact screening scaffold is useful because it treats immune condition as a systems problem shaped by nutrition, barrier integrity, pathogen pressure, pollution, heat stress, and inflammatory burden rather than as a single measurement. A production workflow could add real biomarkers, uncertainty intervals, field metadata, tissue-specific measurements, microbiome data, or longitudinal monitoring.
These examples remain compact enough for an article, but they point toward the kinds of workflows scientists actually use: coupled differential reasoning, threshold effects, scenario comparison, ecological stress screening, and explicit trade-offs between protection and injury.
GitHub repository
The article body includes compact R and Python examples so the biological and scientific argument remains readable. The full repository expands those examples into a broader computational immunology workflow, including host–pathogen dynamics, immune activation and clearance, inflammatory damage, repair and resolution, threshold screening, immune-condition scoring, SQL provenance structures, reproducible data files, and full-stack scientific-computing examples across Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, and notebooks.
Limits, complexity, and modern immunological thinking
Immunology is foundational, but it should not be oversimplified into a simple war metaphor. Immune systems do not merely destroy foreign matter. They classify, regulate, tolerate, remember, repair, coordinate, and sometimes injure. The line between defense and pathology is often thin, and innate and adaptive defenses are integrated, context-dependent, spatially organized, historically shaped, and regulated rather than mechanically uniform.
This matters because not all inflammation is protective, not all tolerance is beneficial, and not all immune memory is equally effective. Autoimmunity, allergy, chronic inflammation, immune exhaustion, immune evasion, immunodeficiency, cancer immune escape, ecological stress, microbiome disruption, aging, and host variation all show that defense is complex and often imperfect.
Models are useful because they clarify assumptions, expose trade-offs, and make immune dynamics testable. But a host–pathogen equation is not a full organism, a threshold score is not a clinical diagnosis, and a simulated inflammatory burden is not a tissue. Quantitative tools are strongest when they support biological interpretation rather than replacing it.
For research biologists, this means models are most useful when they clarify organization and trade-off rather than pretending to eliminate uncertainty, context, or biological complexity.
Why this matters for scientific work
Immunology matters across medicine, microbiology, physiology, ecology, conservation biology, restoration ecology, plant science, animal biology, marine biology, freshwater biology, soil biology, agroecology, forestry, disease ecology, environmental health, biotechnology, and computational biology because biological defense is one of the basic conditions of life. For immunologists, the value is direct: immune systems show how recognition, activation, memory, and regulation preserve organismal integrity. For microbiologists, immunology explains how hosts and microbes negotiate coexistence, conflict, and disease. For physiologists, immunity is a regulatory system distributed across tissues and organs.
For ecologists and conservation biologists, immunology explains how stress, climate change, pollution, nutrition, habitat alteration, and pathogen exposure affect survival and population resilience. For plant scientists and agroecologists, defense systems help explain crop vulnerability, disease resistance, symbiosis, and soil-plant-microbe interaction. For marine and freshwater scientists, immunity helps interpret coral stress, fish disease, amphibian decline, pathogen transmission, and immune compromise under changing aquatic conditions.
For computational readers, immunology provides one of biology’s richest domains for systems modeling, differential equations, agent-based modeling, network analysis, single-cell data, transcriptomics, immune repertoire analysis, threshold screening, and scenario simulation. But its computational treatment remains strongest when tied to tissue biology, organismal physiology, ecology, and empirical validation.
Immunology is therefore more than the study of immune cells. It is the study of how life defends, remembers, tolerates, repairs, and survives.
Conclusion
Immunology and biological defense show that life persists through more than growth, metabolism, development, and regulation alone. Organisms must detect danger, coordinate response, contain infection, regulate inflammation, distinguish self from threat, tolerate what should not be attacked, repair damage, and remember previous encounters. These processes link cells, tissues, organs, microbes, environments, and populations into dynamic systems of protection and controlled risk.
To understand immunology is therefore to understand one of biology’s deepest regulatory achievements: the capacity of living systems to preserve integrity without destroying themselves in the process. Defense is not merely attack. It is recognition, classification, activation, restraint, memory, repair, and ecological negotiation.
Immunology is thus more than the study of immune cells. It is one of the principal ways biology explains how life defends itself at all.
Related articles
- Biology
- Physiology and the Regulation of Living Systems
- Microbiology and the Hidden Majority of Life
- Animal Biology and the Organization of Complex Life
- Plant Biology and the Life of Primary Producers
- Coevolution, Symbiosis, and the Dynamics of Mutual Change
- Behavior, Communication, and Biological Strategy
- Neurobiology and the Organization of Living Response
- Metabolism, Energy, and Biological Function
- Ecology and the Interdependence of Life
Further reading
- Janeway, C.A. Jr, Travers, P., Walport, M. and Shlomchik, M.J. (2001) Immunobiology: The Immune System in Health and Disease. 5th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10757/
- Janeway, C.A. Jr et al. (2001) ‘Principles of Innate and Adaptive Immunity’, in Immunobiology. 5th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK27090/
- Janeway, C.A. Jr et al. (2001) ‘Innate Immunity’, in Immunobiology. 5th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10769/
- Vaillant, A.A.J., Qurie, A. and Hsu, C.W. (2024) ‘Physiology, Immune Response’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK539801/
- Hannoodee, S. and Nasuruddin, D.N. (2024) ‘Acute Inflammatory Response’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK556083/
- Pahwa, R., Goyal, A. and Jialal, I. (2023) ‘Chronic Inflammation’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK493173/
- Medzhitov, R. and Janeway, C.A. Jr (1998) ‘Innate immune recognition and control of adaptive immune responses’, Seminars in Immunology, 10(5), pp. 351–353. Available at: https://pubmed.ncbi.nlm.nih.gov/9799709/
- Janeway, C.A. Jr and Medzhitov, R. (2002) ‘Innate immune recognition’, Annual Review of Immunology, 20, pp. 197–216. Available at: https://pubmed.ncbi.nlm.nih.gov/11861602/
- Romanovsky, A.A. (2007) Toll-like Receptors: Sensors That Detect Infection. Nature Education Scitable. Available at: https://www.nature.com/scitable/topicpage/toll-like-receptors-sensors-that-detect-infection-14396559/
- Shinde, S.B. et al. (2018) ‘Review of the systems biology of the immune system using agent-based models’, Wiley Interdisciplinary Reviews: Systems Biology and Medicine. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC8687247/
- Nature Education Scitable (n.d.) Stress and Animal Health: Physiological Mechanisms and Ecological Consequences. Available at: https://www.nature.com/scitable/knowledge/library/stress-and-animal-health-physiological-mechanisms-and-23672697/
- Nature Education Scitable (n.d.) Host Response to the Dengue Virus. Available at: https://www.nature.com/scitable/topicpage/host-response-to-the-dengue-virus-22402106/
References
- Britannica (n.d.) Immune System. Available at: https://www.britannica.com/science/immune-system
- Britannica (n.d.) Immunology. Available at: https://www.britannica.com/science/immunology
- Britannica (n.d.) The Nature of Lymphocytes. Available at: https://www.britannica.com/science/immune-system/The-nature-of-lymphocytes
- Britannica (n.d.) Classes of Immunoglobulins. Available at: https://www.britannica.com/science/immune-system/Classes-of-immunoglobulins
- Hannoodee, S. and Nasuruddin, D.N. (2024) ‘Acute Inflammatory Response’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK556083/
- Janeway, C.A. Jr, Travers, P., Walport, M. and Shlomchik, M.J. (2001) Immunobiology: The Immune System in Health and Disease. 5th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10757/
- Janeway, C.A. Jr et al. (2001) ‘Basic Concepts in Immunology’, in Immunobiology. 5th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10779/
- Janeway, C.A. Jr et al. (2001) ‘Innate Immunity’, in Immunobiology. 5th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10769/
- Janeway, C.A. Jr et al. (2001) ‘Principles of Innate and Adaptive Immunity’, in Immunobiology. 5th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK27090/
- Janeway, C.A. Jr and Medzhitov, R. (2002) ‘Innate immune recognition’, Annual Review of Immunology, 20, pp. 197–216. Available at: https://pubmed.ncbi.nlm.nih.gov/11861602/
- Libretti, S. and Puckett, Y. (2025) ‘Physiology, Homeostasis’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK559138/
- Medzhitov, R. and Janeway, C.A. Jr (1998) ‘Innate immune recognition and control of adaptive immune responses’, Seminars in Immunology, 10(5), pp. 351–353. Available at: https://pubmed.ncbi.nlm.nih.gov/9799709/
- Nature Education Scitable (n.d.) Host Response to the Dengue Virus. Available at: https://www.nature.com/scitable/topicpage/host-response-to-the-dengue-virus-22402106/
- Nature Education Scitable (n.d.) Stress and Animal Health: Physiological Mechanisms and Ecological Consequences. Available at: https://www.nature.com/scitable/knowledge/library/stress-and-animal-health-physiological-mechanisms-and-23672697/
- Pahwa, R., Goyal, A. and Jialal, I. (2023) ‘Chronic Inflammation’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK493173/
- Romanovsky, A.A. (2007) Toll-like Receptors: Sensors That Detect Infection. Nature Education Scitable. Available at: https://www.nature.com/scitable/topicpage/toll-like-receptors-sensors-that-detect-infection-14396559/
- Shinde, S.B. et al. (2018) ‘Review of the systems biology of the immune system using agent-based models’, Wiley Interdisciplinary Reviews: Systems Biology and Medicine. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC8687247/
- Vaillant, A.A.J., Qurie, A. and Hsu, C.W. (2024) ‘Physiology, Immune Response’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK539801/