
Last Updated May 28, 2026
Physiology and the regulation of living systems examine how organisms maintain functional order through the coordinated control of energy, matter, temperature, water, ions, gases, nutrients, signaling, internal balance, stress response, and adaptive capacity across cells, tissues, organs, and whole organisms. Physiology is central to biology because life does not persist through structure alone. Living systems must regulate themselves continuously in the face of environmental fluctuation, metabolic demand, developmental change, injury, stress, reproduction, disease, and ecological constraint. To study physiology is to study life as regulated activity: the dynamic maintenance of viable conditions under changing internal and external pressures.
Physiology is therefore one of biology’s great integrative sciences. It connects molecular transport to cellular function, tissue specialization to organ performance, organ systems to whole-organism regulation, environmental stress to adaptive response, and computational modeling to the formal analysis of feedback, thresholds, flows, balances, and control. A living system is not merely a collection of parts. It is a coordinated system of exchange, regulation, correction, signaling, and constraint.
Main Library
Publications
Article Map
Biology
Related Topic
Chemistry
Related Topic
Environmental Science
Related Topic
Mathematical Modeling
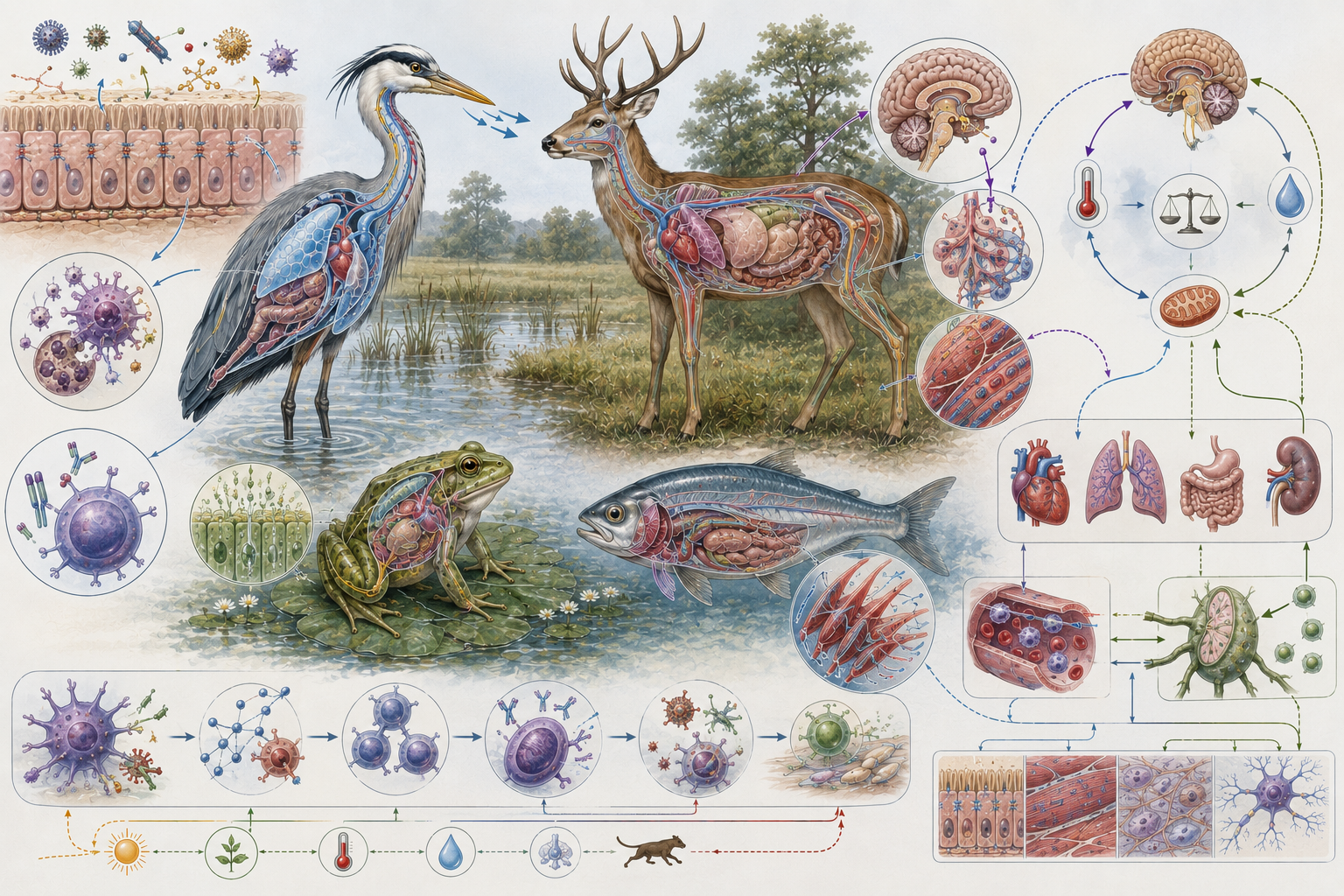
This article develops physiology as a scale-spanning framework for understanding the regulation of living systems. It examines homeostasis, feedback control, internal stability, organ-system coordination, energy and matter balance, water and ion regulation, pH control, thermoregulation, gas exchange, endocrine signaling, neural coordination, immune regulation, plant physiology, animal physiology, microbial physiology, ecological physiology, disease, stress, conservation physiology, systems biology, omics, and computational modeling.
The article is written for physiologists, ecologists, marine biologists, freshwater scientists, medical and environmental-health readers, computational biology readers, biotech readers, biodiversity experts, conservation planners, plant scientists, animal biologists, microbiologists, systems biologists, and research biologists who need a rigorous account of how regulation operates across scales, taxa, habitats, and changing conditions.
The article also extends physiology into quantitative and computational biology through balance equations, negative-feedback models, coupled signaling and effector systems, regulatory performance scoring, stress scenarios, R workflows, Python workflows, SQL provenance structures, and a linked full-stack GitHub repository containing Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, notebooks, data files, and reproducibility documentation.
What physiology studies
Physiology is the study of function in living systems. It asks how organisms work: how they circulate materials, exchange gases, regulate temperature, maintain water and ion balance, process nutrients, coordinate organs, respond to stress, repair disruption, and preserve internal viability across time. Physiology is therefore not merely a catalog of organs or mechanisms. It is the study of dynamic living order.
This matters because life depends on continuous activity rather than static structure. A cell membrane, nephron, leaf, heart, gill, root, endocrine gland, epithelial surface, microbial transporter, or vascular tissue is biologically meaningful not only because it exists, but because it participates in regulated function. Physiology provides the explanatory framework for understanding how those parts operate as living systems rather than as anatomical objects.
Physiology also clarifies that function is inseparable from constraint. Organisms do not simply perform processes. They perform them within ranges: temperature ranges, oxygen ranges, osmotic ranges, pH ranges, pressure ranges, nutrient ranges, hydration ranges, and energetic limits. Physiological function is therefore the continuous work of maintaining viability under constraint.
For research biologists, physiology belongs near the center of biology because it links structure to process, process to regulation, and regulation to survival under real environmental, developmental, and ecological conditions.
Homeostasis and the problem of living order
One of physiology’s core concepts is homeostasis: the capacity of living systems to maintain relatively stable internal conditions while adjusting to changing environments. Homeostasis is best understood not as static constancy but as constrained variability. Internal temperature, blood glucose, osmotic state, oxygen availability, pH, ion concentrations, hydration status, and stress mediators all fluctuate, yet not without limit.
This matters because living systems must keep many variables within workable ranges rather than at one perfectly fixed point. Stability in physiology is therefore a dynamic equilibrium, not a frozen state. Organisms persist by regulating ongoing change, not by escaping change altogether.
Homeostasis also reveals why physiology is fundamentally active. Maintaining order requires energy, sensing, correction, transport, communication, and feedback. A body must detect deviation, signal that deviation, mobilize effectors, and alter flow, uptake, release, behavior, or metabolism. Homeostasis is not passive balance. It is controlled adjustment.
For research biologists, homeostasis remains powerful because it offers a unifying concept while also inviting refinement. Different organisms regulate different variables, at different time scales, with different tolerances, and often through multiple overlapping systems rather than one simple control loop.
Feedback control and the logic of regulation
Physiological regulation is often organized through feedback. Negative feedback counteracts deviation from a functional range, while positive feedback can amplify a process when rapid completion is biologically useful. Negative feedback is especially important in homeostasis because it helps stabilize variables such as temperature, glucose, pH, blood pressure, oxygen supply, and fluid balance.
This matters because physiology is not only chemistry happening inside a body. It is organized control. Sensors detect change, integrative centers interpret that change, and effectors act to alter system state. Thermoregulation, endocrine control, ventilation, renal adjustment, acid–base compensation, blood-pressure regulation, osmoregulation, and many immune and metabolic processes all follow this broader logic.
Feedback systems also have limits. Delays, overshoot, weak sensing, exhausted effectors, conflicting demands, or external stress can produce oscillation, instability, or failure. A regulatory system may work under ordinary conditions and fail under heat, dehydration, hypoxia, infection, toxic exposure, or metabolic overload.
For research biologists, feedback is especially useful because it connects physiology to control theory, systems biology, and computational modeling. Regulation becomes analyzable as a problem of sensing, deviation, correction, delay, capacity, and constraint.
Cells, tissues, organs, and levels of physiological integration
Physiology operates across nested levels of organization. Cells regulate membrane transport, energy use, signaling, protein turnover, osmotic balance, and internal chemistry. Tissues coordinate specialized cellular functions. Organs integrate those tissues into functional units such as lungs, kidneys, roots, leaves, hearts, livers, gills, muscles, glands, or vascular tissues. Organ systems coordinate multiple organs into still larger regulatory wholes.
This matters because no major physiological problem is solved at only one level. Gas exchange depends on molecular diffusion, epithelial surfaces, vascular transport, ventilation, circulation, and whole-organism demand. Glucose balance depends on digestion, endocrine signaling, liver metabolism, tissue uptake, cellular transport, and neural control. Water regulation depends on membrane transport, blood volume, renal function, hormonal signaling, behavior, and environmental exposure.
Physiological integration also means that local events can have systemic consequences. A kidney problem can alter blood pressure, ion balance, acid–base state, and cardiovascular load. A respiratory problem can alter oxygen delivery, carbon dioxide removal, pH, and cellular metabolism. A plant stomatal response can alter water loss, carbon gain, temperature, and growth.
For research biologists, physiology is therefore a science of integration. The regulation of living systems depends on interactions across scales, not isolated parts acting alone. Multi-scale explanation is not optional here. It is built into the subject.
Energy, matter, and the economy of living systems
All physiology depends on the managed movement of energy and matter. Organisms must acquire, transform, distribute, store, and expend energy while also maintaining the transport and balance of water, nutrients, gases, ions, heat, wastes, and signals. Physiological function is therefore inseparable from biological economy: what enters, what is transformed, what is retained, what is lost, and what must be continuously replaced.
This matters because regulation is not abstract. It is constrained by metabolic cost, environmental availability, and structural capacity. A body cannot regulate glucose without acquiring or mobilizing substrates. It cannot regulate temperature without balancing heat generation and loss. It cannot regulate ion state without active transport, membrane selectivity, and water balance. It cannot maintain stress response indefinitely without energetic and tissue consequences.
Physiology therefore reveals that life is budgeted. Growth, maintenance, repair, reproduction, immune defense, thermoregulation, movement, detoxification, osmotic control, and stress adaptation compete for limited energy and matter. A regulatory success in one domain may impose costs elsewhere.
For research biologists, this is one of physiology’s deepest lessons: every regulatory achievement has an energetic and material cost. Living systems are not only organized. They are constrained economies of function.
Water, ions, pH, and internal chemical balance
Water balance, ion concentrations, and acid–base regulation are among physiology’s most fundamental problems because life depends on narrow chemical ranges. Many enzymes, membranes, transporters, organelles, signaling systems, and macromolecular assemblies function only within constrained ionic and pH conditions. Too much or too little sodium, potassium, calcium, chloride, hydrogen ion, bicarbonate, or water can destabilize function. Too much acidity or alkalinity can impair enzyme performance, oxygen transport, membrane excitability, cellular signaling, and viability.
This matters because cells are chemical systems before they are anything else. Chemical regulation is therefore not a background feature of physiology. It is one of its foundations. Osmotic pressure, buffering, ion gradients, membrane potentials, fluid compartmentation, active transport, and epithelial exchange all help determine whether life can continue.
These processes are also environmentally situated. A freshwater fish faces different osmotic challenges from a marine fish. A desert plant faces different water-balance problems from a wetland plant. A soil microbe faces fluctuating moisture and chemical gradients. An animal under dehydration, heat stress, or kidney dysfunction faces immediate regulatory strain.
For research biologists, internal chemical balance is especially important because it links molecular biophysics, cell biology, whole-organism physiology, and environmental stress in one shared explanatory framework.
Thermoregulation, gas exchange, and environmental constraint
Physiology also depends on the regulation of temperature and gases. Internal temperature affects enzyme kinetics, membrane properties, muscle performance, developmental timing, neural function, immune response, and energy metabolism. Gas exchange determines whether oxygen can support aerobic function and whether carbon dioxide can be removed before acid–base disturbance intensifies. Lungs, gills, tracheae, leaves, stomata, vascular systems, and microbial membranes all solve different versions of this problem.
This matters because organisms exist within physical environments that can destabilize internal function. Heat, cold, oxygen limitation, carbon dioxide accumulation, salinity, drought, pressure, altitude, turbidity, hypoxia, and water availability all impose physiological constraints. Gas exchange and thermal regulation therefore belong to a larger problem of staying chemically and thermally viable under changing external conditions.
Thermoregulation is especially useful as a model of physiological integration. It may involve behavior, circulation, sweating, panting, shivering, insulation, evaporative cooling, metabolic heat production, vascular adjustment, posture, dormancy, leaf orientation, or habitat selection. Temperature regulation is not only a biochemical problem. It is a whole-organism and ecological problem.
For research biologists, physiology becomes environmental biology from the inside: the study of how organisms remain functional within material limits rather than how they merely occupy environments in descriptive terms.
Endocrine, neural, and immune coordination
Regulation in complex organisms depends strongly on communication systems. Hormones coordinate slower systemic responses across tissues. Nervous systems support rapid signaling, integration, and behavioral adjustment. Immune systems monitor and respond to microbial intrusion, tissue damage, inflammation, and altered internal states. Stress responses, developmental transitions, circadian timing, reproduction, inflammation, metabolism, and repair all depend on these interacting communication systems.
This matters because physiology is not only a set of parallel subsystems. It is a coordinated whole in which internal states are sensed, communicated, interpreted, and adjusted through interacting networks. Glucose control, reproduction, thermoregulation, inflammation, circadian timing, growth, immune defense, and stress response all depend on these integrative signaling systems.
The endocrine, neural, and immune systems also interact with one another. Stress hormones affect immune function. Inflammation affects neural and endocrine regulation. Neural control influences heart rate, digestion, breathing, blood pressure, and thermoregulation. Hormones alter metabolism, development, reproduction, and behavior. Regulation is therefore distributed rather than confined to one commanding system.
For research biologists, this makes physiology one of the clearest domains in which signal and substance meet. Regulation requires both material exchange and information flow.
Physiology in plants, animals, and microbes
Although physiology is often taught through animal examples, all major forms of life solve regulatory problems. Plants regulate water, stomatal conductance, nutrient uptake, hormone signaling, osmotic stress, light response, growth allocation, carbon balance, and defense. Animals regulate circulation, temperature, glucose, gases, fluid balance, internal organs, locomotion, reproduction, and stress response. Microbes regulate membrane transport, metabolism, stress response, quorum behavior, osmotic adaptation, dormancy, growth state, and environmental sensing.
This matters because physiology is not restricted to one kingdom. It is a general biological problem of how living systems maintain function under internal and external constraint. The mechanisms differ, but the logic of regulation persists across life.
Comparative physiology also reveals that there is no single universal solution to regulation. Some organisms conform to environmental conditions more than they regulate them. Others regulate tightly. Some tolerate extreme variation. Others survive only within narrow ranges. Some shift metabolism, enter dormancy, accumulate solutes, alter membrane composition, or use behavioral avoidance.
For research biologists, comparative physiology is especially valuable because it reveals how deeply conserved the problem of regulation is, even where the mechanistic solutions vary dramatically.
Ecology and the physiological limits of life
Ecology is partly physiology written across environments. Organisms maximize performance within particular environmental ranges and can persist only within broader tolerance limits. Distribution, abundance, competition, phenology, disease susceptibility, reproduction, and resilience all depend partly on physiological capacity: water balance, temperature tolerance, gas exchange, nutrient uptake, developmental timing, stress response, and metabolic flexibility.
This matters because ecological boundaries are often physiological before they are demographic. An organism occurs where it can maintain regulated function, not merely where it can arrive. Physiological thresholds therefore help determine habitat suitability, range limits, seasonal timing, migration constraints, reproductive windows, and vulnerability under change.
Ecophysiology is especially important under climate change. Heat waves, droughts, hypoxia, salinity shifts, ocean acidification, nutrient imbalance, and altered precipitation regimes are not simply environmental trends. They are regulatory pressures on organisms. Populations may decline not only because habitats disappear, but because the physiological costs of persistence rise beyond tolerable limits.
For research biologists, this is one of the strongest bridges between physiology and ecology. The limits of life in place are often the limits of regulation under environmental constraint.
Marine, freshwater, soil, and terrestrial physiology
Different environments impose different regulatory burdens. Marine organisms must manage salinity, buoyancy, pressure, oxygen gradients, acid–base balance, and temperature variation. Freshwater organisms face osmotic challenges, dissolved oxygen instability, hydrologic fragmentation, turbidity, and flow change. Soil organisms confront low-oxygen microsites, variable moisture, intense chemical gradients, organic matter dynamics, and particulate environments. Terrestrial organisms must regulate desiccation, temperature, gas exchange, posture, exposure, and water conservation in air.
This matters because physiology is always material and situated. The same broad regulatory logic takes different forms in a fish, amphibian, fungus, bacterium, tree, insect, bird, mammal, coral, mollusk, or soil microbe because the environment changes what must be controlled and how.
Marine physiology is increasingly important for understanding warming seas, hypoxia, acidification, and coral bleaching. Freshwater physiology matters for rivers, lakes, wetlands, fish migration, amphibian decline, and dissolved oxygen stress. Soil physiology matters for microbial function, root systems, fungal networks, drought, nutrient cycling, and agroecology. Terrestrial physiology matters for heat stress, water balance, migration, metabolism, and reproductive timing.
For research biologists, environmental situatedness is critical. Warming oceans, drought-stricken soils, salinized fields, fragmented rivers, and oxygen-poor waters are not just environmental descriptors. They are regulatory crises for living systems.
Disease, stress, and the breakdown of regulation
Disease can be understood physiologically as the partial breakdown, overload, or maladaptive redirection of regulation. Compensation is exhausted. Signaling becomes distorted. Internal conditions drift outside viable ranges. Heat stroke, dehydration, acidosis, alkalosis, shock, glucose dysregulation, immune overreaction, respiratory failure, kidney failure, chronic inflammation, endocrine disorder, and autonomic dysfunction can all be understood as failures or overloads of regulated balance rather than as unrelated phenomena.
This matters because physiology helps unify pathology. Organisms must respond continuously to internal and external disturbances to preserve function. When the response is too weak, too delayed, too costly, too prolonged, or itself damaging, breakdown emerges.
Stress is especially important because it reveals the cost of regulation. Acute stress responses may protect function under immediate challenge. Chronic stress can erode immune defense, metabolism, reproduction, growth, neural function, and repair. What is adaptive in one time frame may become damaging in another.
For research biologists, physiology is one of the strongest bridges between normal function and pathological disruption. Disease often clarifies how regulation was working in the first place by showing what happens when it fails.
Conservation, systems thinking, and the regulation of living systems
Physiology is central to conservation and systems-oriented biology because organisms under environmental change survive or fail through their regulatory capacities. Heat tolerance, drought resistance, osmotic flexibility, acid–base compensation, oxygen use, stress signaling, immune function, reproductive timing, metabolic plasticity, and developmental flexibility all influence whether populations persist under changing conditions.
This matters because conservation problems are not only about landscapes or species counts. They are also about physiological viability. A forest exposed to prolonged drought, a coral reef under heat stress, a freshwater fish under hypoxia, a soil microbiome under chemical contamination, or a pollinator under pesticide and thermal stress all face regulatory challenges before they face extinction outright.
Conservation physiology asks how organisms maintain function under real environmental pressures and how those limits shape vulnerability, resilience, and recovery. It connects organismal biology to population viability, habitat quality, climate adaptation, restoration success, and ecosystem function.
For research biologists, physiology helps reveal how environmental crisis becomes biological consequence. It is one of the principal sciences of resilience, stress, and limit.
Systems biology, omics, and computational relevance
Modern physiology increasingly depends on systems biology, multi-omics data, imaging, modeling, sensor networks, and computational integration. Regulation is now studied not only at the level of single organs or variables, but across interacting networks of genes, proteins, metabolites, cells, tissues, organs, behavior, and environmental cues. Physiological states can be inferred from transcriptomic, proteomic, metabolomic, signaling, imaging, time-series, and whole-organism measurements together.
This matters because physiology is inherently multiscale. A homeostatic response may involve transcriptional change, membrane transport, hormonal signaling, tissue remodeling, behavioral adjustment, immune modulation, and ecological consequence all at once. Computational approaches help make such complexity legible.
Systems physiology also makes assumptions explicit. Models can identify variables, rates, delays, thresholds, feedback loops, stressors, and sensitivity points. They can compare weak and strong feedback, delayed response, increased load, compensatory failure, or altered effector capacity. This does not replace biological experiment or field observation. It supports them by clarifying how regulation might work.
For research biologists, physiology sits naturally at the intersection of classical integrative biology and contemporary systems science. It is one of the clearest arenas in which reductionist and integrative approaches must genuinely meet.
Quantitative physiology: mathematics, R, and Python
Physiology is deeply quantitative because regulation often depends on rates, balances, thresholds, delays, capacities, and feedback dynamics. A simple balance model begins by expressing the rate of change of a regulated variable as the difference between inputs and outputs:
\frac{dX}{dt}=I-O
\]
Interpretation: \(X\) is the internal quantity of interest, \(I\) is input, and \(O\) is output. This is useful because many physiological problems reduce, at first approximation, to whether material or energy enters faster than it leaves.
A simple negative-feedback representation can be written as:
\frac{dX}{dt}=-k(X-X^*)
\]
Interpretation: \(X^*\) is the target range or set value, and \(k\) is the strength of correction. This captures the intuition that the farther a variable drifts from its regulated state, the stronger corrective response may become.
For research biologists, however, physiology usually requires more structure than one-variable recovery. A compact coupled formulation for a regulated metabolite, hormone signal, and effector response can be written as:
\frac{dX}{dt}=I-U(H,X)
\]
Interpretation: \(X\) is the regulated variable, and \(U(H,X)\) is hormone-dependent uptake or correction.
\frac{dH}{dt}=a(X-X^*)-bH
\]
Interpretation: \(H\) is a hormonal or signaling intermediate, with \(a\) representing sensing or activation strength and \(b\) representing signal decay.
\frac{dE}{dt}=cH-dE
\]
Interpretation: \(E\) is an effector process, with \(c\) and \(d\) governing effector activation and resolution. The coupled system expresses a central physiological truth: regulation often involves sensing, signaling, and effectors rather than direct automatic correction.
Worked example: fluid balance
Suppose an organism takes in water at 2.5 liters per day and loses water at 2.1 liters per day. Then:
\frac{dX}{dt}=2.5-2.1=0.4
\]
Interpretation: Body water would rise by 0.4 liters per day under the simplified model if no compensatory regulation occurred. Physiology then asks what mechanisms reduce that imbalance before it becomes destabilizing.
R and Python workflows
The following examples are compact article-level workflows. The full GitHub repository expands them into richer multi-language implementations with SQL provenance, validation notes, balance equations, coupled feedback systems, hormonal signaling, effector response, stress scenarios, regulatory performance scoring, thermal and fluid-balance examples, and reproducible computational physiology scaffolding.
R example: coupled homeostatic recovery with hormonal feedback
# Coupled physiological regulation model in R
#
# This compact workflow models:
# - regulated variable X
# - hormonal controller H
# - effector response E
# - uptake or correction as a function of H and X
#
# Euler integration is used for article-scale clarity.
# This is a teaching scaffold, not a calibrated clinical model.
dt <- 0.05
time <- seq(0, 40, by = dt)
# Parameters
X_star <- 5
I_in <- 0.6
a <- 0.9
b <- 0.5
c <- 0.7
d <- 0.4
u0 <- 0.3
u1 <- 0.25
# State variables
X <- numeric(length(time))
H <- numeric(length(time))
E <- numeric(length(time))
# Initial condition: perturbed internal variable
X[1] <- 10
H[1] <- 0
E[1] <- 0
for (t in 2:length(time)) {
uptake <- u0 + u1 * H[t - 1] * X[t - 1]
dX <- I_in - uptake
dH <- a * (X[t - 1] - X_star) - b * H[t - 1]
dE <- c * H[t - 1] - d * E[t - 1]
X[t] <- max(0, X[t - 1] + dX * dt)
H[t] <- max(0, H[t - 1] + dH * dt)
E[t] <- max(0, E[t - 1] + dE * dt)
}
results <- data.frame(
time = time,
regulated_variable = X,
hormonal_signal = H,
effector_response = E
)
matplot(
time,
cbind(X, H, E),
type = "l",
lty = 1,
lwd = 2,
xlab = "Time",
ylab = "State value",
main = "Coupled Physiological Regulation"
)
abline(h = X_star, lty = 2)
legend(
"topright",
legend = c("Regulated variable X", "Hormonal signal H", "Effector E"),
lty = 1,
lwd = 2,
bty = "n"
)
print(tail(round(results, 3), 10))
diagnostics <- data.frame(
peak_X = max(X),
peak_H = max(H),
peak_E = max(E),
final_X = tail(X, 1),
recovery_error = abs(tail(X, 1) - X_star)
)
print(round(diagnostics, 3))This R workflow is more useful than a single exponential return curve because it represents physiology as a coupled control system rather than a one-parameter relaxation process. A research biologist could adapt it for glucose regulation, osmotic balance, thermal compensation, endocrine control, acid–base recovery, stress physiology, or comparative regulatory performance.
Python example: comparative feedback strengths and stress scenarios
import numpy as np
import pandas as pd
def simulate_feedback(
X0=10,
X_star=5,
I_in=0.6,
a=0.9,
b=0.5,
c=0.7,
d=0.4,
u0=0.3,
u1=0.25,
T=40,
dt=0.05
):
"""Simulate a coupled physiological feedback system."""
time = np.arange(0, T + dt, dt)
X = np.zeros(len(time))
H = np.zeros(len(time))
E = np.zeros(len(time))
X[0] = X0
H[0] = 0
E[0] = 0
for t in range(1, len(time)):
uptake = u0 + u1 * H[t - 1] * X[t - 1]
dX = I_in - uptake
dH = a * (X[t - 1] - X_star) - b * H[t - 1]
dE = c * H[t - 1] - d * E[t - 1]
X[t] = max(0, X[t - 1] + dX * dt)
H[t] = max(0, H[t - 1] + dH * dt)
E[t] = max(0, E[t - 1] + dE * dt)
return {
"final_X": X[-1],
"peak_X": X.max(),
"peak_H": H.max(),
"peak_E": E.max(),
"recovery_error": abs(X[-1] - X_star)
}
scenarios = {
"weak_feedback": {"a": 0.5, "b": 0.5, "u1": 0.15},
"moderate_feedback": {"a": 0.9, "b": 0.5, "u1": 0.25},
"strong_feedback": {"a": 1.3, "b": 0.6, "u1": 0.35},
"stress_high_input": {"a": 0.9, "b": 0.5, "u1": 0.25, "I_in": 0.9},
"weak_effector": {"a": 0.9, "b": 0.5, "u1": 0.12}
}
rows = []
for name, params in scenarios.items():
result = simulate_feedback(**params)
result["scenario"] = name
rows.append(result)
df = pd.DataFrame(rows)
df["regulatory_class"] = np.where(
df["recovery_error"] < 0.5,
"well-regulated",
np.where(
df["recovery_error"] < 1.5,
"strained",
"poorly-regulated"
)
)
print(df.round(3))This Python workflow is more useful because it compares multiple regulatory regimes rather than plotting one fixed recovery process. It allows the reader to examine how stronger or weaker feedback, higher input stress, and weaker effector capacity alter control quality, signal burden, and final recovery error. That makes it adaptable to comparative physiology, stress biology, ecophysiology, conservation physiology, and systems-biology teaching.
Python example: physiological stress screening across habitats
import numpy as np
import pandas as pd
# Synthetic habitat-level physiological stress indicators.
# Values are scaled between 0 and 1 for article clarity.
habitats = pd.DataFrame({
"habitat": ["reef", "wetland", "forest", "grassland", "river"],
"thermal_stress": [0.82, 0.44, 0.38, 0.61, 0.49],
"water_stress": [0.30, 0.28, 0.47, 0.68, 0.35],
"oxygen_stress": [0.52, 0.58, 0.25, 0.31, 0.66],
"salinity_or_ion_stress": [0.64, 0.41, 0.22, 0.30, 0.55],
"nutrient_constraint": [0.46, 0.38, 0.42, 0.57, 0.50],
"regulatory_plasticity": [0.48, 0.62, 0.70, 0.55, 0.58]
})
habitats["physiological_stress_index"] = (
0.22 * habitats["thermal_stress"]
+ 0.20 * habitats["water_stress"]
+ 0.18 * habitats["oxygen_stress"]
+ 0.15 * habitats["salinity_or_ion_stress"]
+ 0.15 * habitats["nutrient_constraint"]
- 0.20 * habitats["regulatory_plasticity"]
)
habitats["risk_class"] = np.where(
habitats["physiological_stress_index"] > 0.45,
"high physiological strain",
np.where(
habitats["physiological_stress_index"] > 0.30,
"moderate strain",
"relatively buffered"
)
)
print(habitats.round(3).to_string(index=False))This compact screening scaffold is useful because it treats physiological viability as a multi-factor systems problem rather than a single stressor. A production workflow could add measured thermal limits, dissolved oxygen, osmotic parameters, pH, organismal performance curves, uncertainty intervals, longitudinal field data, or species-specific tolerance thresholds.
These examples remain compact enough for an article, but they point toward the kinds of workflows scientists actually use: coupled differential reasoning, control-logic comparison, stress scenarios, performance classification, ecological stress screening, and explicit tracking of regulatory error rather than one illustrative decay curve alone.
GitHub repository
The article body includes compact R and Python examples so the biological and scientific argument remains readable. The full repository expands those examples into a broader computational physiology workflow, including balance equations, coupled feedback systems, hormonal signaling, effector response, stress scenarios, regulatory performance scoring, thermal and fluid-balance examples, SQL provenance structures, reproducible data files, and full-stack scientific-computing examples across Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, and notebooks.
Limits, complexity, and modern physiological thinking
Physiology is foundational, but it should not be oversimplified. Homeostasis is a powerful unifying concept, yet real regulation is often multilevel, dynamic, context-dependent, historically evolved, and not reducible to one set-point metaphor. Internal stability is maintained through interacting systems, imperfect compensation, trade-offs, oscillation, feedforward signals, behavioral adjustment, and sometimes adaptive shifts rather than rigid constancy.
This matters because physiology is not a static engineering diagram. It is a living, path-dependent, environmentally situated process. Not all organisms regulate the same variables in the same way. Not all disturbances are fully compensated. Not all stable states are healthy ones. A system can be stable but maladaptive, compensated but strained, or resilient only within a narrow range.
Models are useful because they clarify assumptions, expose feedback structure, and make regulatory performance testable. But a feedback equation is not a whole organism, a set point is not a complete physiology, and a recovery error is not the same as biological health. Quantitative tools are strongest when they support biological interpretation rather than replacing it.
For research biologists, modern physiological thinking is strongest when it preserves the centrality of regulation while also recognizing complexity, multiscale interaction, adaptive limitation, and environmental context.
Why this matters for scientific work
Physiology matters across medicine, ecology, conservation biology, restoration ecology, plant science, animal biology, microbiology, marine biology, freshwater biology, soil biology, environmental health, biotechnology, disease ecology, and computational biology because regulation is one of the basic conditions of life. For physiologists, the value is direct: physiology explains how living systems maintain function through feedback, exchange, signaling, and constraint. For medical and environmental-health readers, physiology clarifies how disease, exposure, stress, heat, dehydration, hypoxia, toxic substances, and immune disruption become functional breakdowns.
For ecologists and conservation biologists, physiology explains why organisms persist in some habitats and fail in others. Temperature tolerance, water balance, oxygen availability, salinity, acid–base compensation, nutrient uptake, and stress response can determine species distributions, reproductive windows, migration patterns, disease susceptibility, and resilience under climate change. For plant scientists and agroecologists, physiology helps explain drought tolerance, stomatal control, nutrient uptake, salinity stress, growth allocation, and yield vulnerability.
For computational readers, physiology provides one of biology’s richest domains for systems modeling, differential equations, control theory, time-series analysis, threshold screening, parameter sensitivity, digital twins, and reproducible simulation. But its computational treatment remains strongest when tied to experimental biology, field observation, organismal function, and environmental context.
Physiology is therefore more than the study of how bodies work. It is the study of how living systems remain organized under pressure.
Conclusion
Physiology and the regulation of living systems show that life persists through coordinated function rather than structure alone. Organisms regulate temperature, water, ions, gases, nutrients, stress responses, signaling systems, and internal balance through dynamic systems of feedback, exchange, correction, and integration across cells, tissues, organs, and whole bodies.
To understand physiology is therefore to understand one of biology’s deepest explanatory domains: how living systems remain viable under continuous internal and external challenge. Regulation is not a minor maintenance function. It is the active work of staying alive under constraint.
Physiology is thus more than the study of how bodies work. It is one of the principal ways biology explains how life remains organized at all.
Related articles
- Biology
- Metabolism, Energy, and Biological Function
- Development, Differentiation, and the Making of Organisms
- Animal Biology and the Organization of Complex Life
- Plant Biology and the Life of Primary Producers
- Microbiology and the Hidden Majority of Life
- Immunology and Biological Defense
- Neurobiology and the Organization of Living Response
- Ecology and the Interdependence of Life
- Biogeochemical Cycles and the Conditions of Habitability
- Population Dynamics and Ecological Modeling
Further reading
- Billman, G.E. (2020) ‘Homeostasis: The underappreciated and far too often ignored central organizing principle of physiology’, Frontiers in Physiology, 11, article 200. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC7076167/
- Libretti, S. and Puckett, Y. (2025) ‘Physiology, Homeostasis’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK559138/
- Goldstein, D.S. (2019) ‘How does homeostasis happen? Integrative physiological, systems biological, and evolutionary perspectives’, Autonomic Neuroscience, 218, pp. 44–52. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC6483214/
- Modell, H. et al. (2015) ‘A physiologist’s view of homeostasis’, Advances in Physiology Education, 39(4), pp. 259–266. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC4669363/
- De Luca Jr, L.A. et al. (2014) Homeostasis and Body Fluid Regulation. NCBI Bookshelf. Available at: https://www.ncbi.nlm.nih.gov/books/NBK200958/
- Hopkins, E., Sharma, S. and Dhamoon, A.S. (2022) ‘Physiology, Acid Base Balance’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK507807/
- Osilla, E.V. et al. (2023) ‘Physiology, Temperature Regulation’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK507838/
- Romanovsky, A.A. (2018) ‘The thermoregulation system and how it works’, Handbook of Clinical Neurology, 156, pp. 3–43. Available at: https://pubmed.ncbi.nlm.nih.gov/30454596/
- Romanovsky, A.A. (2007) Homeostatic Processes for Thermoregulation. Nature Education Scitable. Available at: https://www.nature.com/scitable/knowledge/library/homeostatic-processes-for-thermoregulation-23592046/
- Sunday, J.M., Bates, A.E. and Dulvy, N.K. (2012) ‘Thermal tolerance and the global redistribution of animals’, Nature Climate Change, 2, pp. 686–690. Available at: https://doi.org/10.1038/nclimate1539
- Somero, G.N. (2010) ‘The physiology of climate change: How potentials for acclimatization and genetic adaptation will determine “winners” and “losers”’, Journal of Experimental Biology, 213(6), pp. 912–920. Available at: https://doi.org/10.1242/jeb.037473
- Carey, N. and Sigwart, J.D. (2014) ‘Size matters: Plasticity in metabolic scaling shows body-size may modulate responses to climate change’, Biology Letters, 10(7), 20140408. Available at: https://doi.org/10.1098/rsbl.2014.0408
References
- Billman, G.E. (2020) ‘Homeostasis: The underappreciated and far too often ignored central organizing principle of physiology’, Frontiers in Physiology, 11, article 200. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC7076167/
- Britannica (2026) Homeostasis. Available at: https://www.britannica.com/science/homeostasis
- Britannica (2026) Physiology: Regulation. Available at: https://www.britannica.com/science/physiology/Regulation
- Carey, N. and Sigwart, J.D. (2014) ‘Size matters: Plasticity in metabolic scaling shows body-size may modulate responses to climate change’, Biology Letters, 10(7), 20140408. Available at: https://doi.org/10.1098/rsbl.2014.0408
- De Luca Jr, L.A. et al. (2014) Homeostasis and Body Fluid Regulation. NCBI Bookshelf. Available at: https://www.ncbi.nlm.nih.gov/books/NBK200958/
- Goldstein, D.S. (2019) ‘How does homeostasis happen? Integrative physiological, systems biological, and evolutionary perspectives’, Autonomic Neuroscience, 218, pp. 44–52. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC6483214/
- Hopkins, E., Sharma, S. and Dhamoon, A.S. (2022) ‘Physiology, Acid Base Balance’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK507807/
- Libretti, S. and Puckett, Y. (2025) ‘Physiology, Homeostasis’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK559138/
- Modell, H., Cliff, W., Michael, J., McFarland, J., Wenderoth, M.P. and Wright, A. (2015) ‘A physiologist’s view of homeostasis’, Advances in Physiology Education, 39(4), pp. 259–266. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC4669363/
- Osilla, E.V. et al. (2023) ‘Physiology, Temperature Regulation’, StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK507838/
- Romanovsky, A.A. (2007) Homeostatic Processes for Thermoregulation. Nature Education Scitable. Available at: https://www.nature.com/scitable/knowledge/library/homeostatic-processes-for-thermoregulation-23592046/
- Romanovsky, A.A. (2018) ‘The thermoregulation system and how it works’, Handbook of Clinical Neurology, 156, pp. 3–43. Available at: https://pubmed.ncbi.nlm.nih.gov/30454596/
- Somero, G.N. (2010) ‘The physiology of climate change: How potentials for acclimatization and genetic adaptation will determine “winners” and “losers”’, Journal of Experimental Biology, 213(6), pp. 912–920. Available at: https://doi.org/10.1242/jeb.037473
- Sunday, J.M., Bates, A.E. and Dulvy, N.K. (2012) ‘Thermal tolerance and the global redistribution of animals’, Nature Climate Change, 2, pp. 686–690. Available at: https://doi.org/10.1038/nclimate1539