
Last Updated May 28, 2026
Cell signaling, communication, and biological coordination examine how living systems detect information, transmit signals, interpret changing conditions, and coordinate activity across molecules, cells, tissues, organisms, populations, and environments. Signaling is one of the central principles of biology because no living system can remain organized without the ability to sense conditions, respond selectively, and coordinate internal and external processes across time. Cells must detect nutrients, pathogens, damage, developmental cues, neighbors, hormones, metabolites, mechanical force, light, toxins, osmotic stress, and changing ecological conditions. Biological organization is therefore not only a matter of structure, metabolism, or genetic information. It is also a matter of regulated communication.
This article develops Cell Signaling, Communication, and Biological Coordination as a foundational article within the Biology knowledge series. It treats signaling not merely as a list of named pathways, but as one of biology’s major coordination systems: the framework through which molecular detection becomes cellular response, cellular response becomes tissue-level organization, and organismal response becomes ecological and evolutionary consequence. Receptors, ligands, second messengers, phosphorylation cascades, calcium transients, feedback loops, quorum signals, hormones, cytokines, neurotransmitters, plant signaling molecules, and microbial communication systems all express the same deeper biological problem: how living systems convert information into coordinated action.
Main Library
Publications
Article Map
Biology
Related Topic
Chemistry
Related Topic
Earth Science
Related Topic
Environmental Science
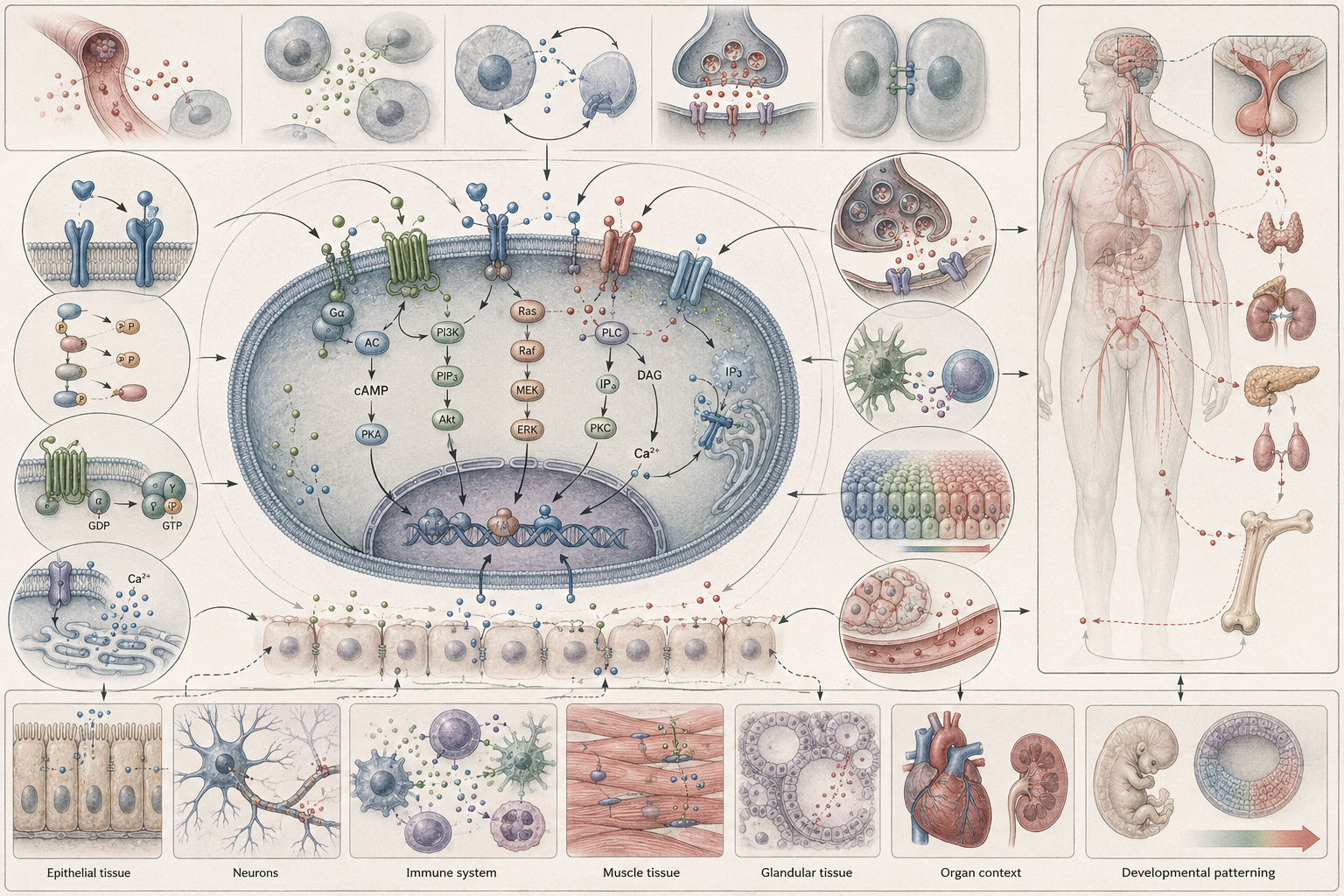
The article develops cell signaling, communication, and biological coordination as a scale-spanning framework for understanding signaling molecules, receptors, signal transduction, GPCRs, receptor tyrosine kinases, intracellular receptors, ion-channel-linked receptors, second messengers, calcium signaling, cAMP and cGMP signaling, kinase cascades, pathway cross-talk, feedback control, development, physiology, immunity, microbial communication, plant signaling, ecological response, disease, biotechnology, and systems biology.
The article also extends signaling into quantitative and computational biology through receptor-occupancy models, Hill response curves, signaling half-life estimation, second-messenger decay, pulse-driven pathway dynamics, negative-feedback systems, calcium/cAMP-style time series, quorum-sensing threshold logic, pathway activation scoring, R workflows, Python workflows, SQL provenance structures, and a linked full-stack GitHub repository containing Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, notebooks, data files, and reproducibility documentation.
What cell signaling is
Cell signaling is the organized process through which cells detect information, transmit it through molecular pathways, and generate specific responses. A signal may originate outside a cell, inside a cell, or across cell-cell contact. It may take the form of a hormone, neurotransmitter, growth factor, metabolite, ion gradient, mechanical force, pathogen-derived molecule, light cue, nutrient state, toxin, osmotic change, or stress condition. What makes signaling biologically important is not simply that information exists, but that cells can detect it selectively and act on it in ways that preserve function and coordination.
Signaling therefore sits at the heart of biological organization. Cells must know whether nutrients are available, whether DNA is damaged, whether neighbors are present, whether pathogens are invading, whether development should proceed, whether tissues require repair, whether stress responses should be activated, and whether environmental conditions are shifting. None of these responses can be left to chemical accident. They depend on pathways that connect detection to regulated action.
This makes cell signaling one of the major bridges between molecular biology and systems biology. Information enters through receptors or intracellular sensors, is distributed through pathways, amplified or dampened by intracellular components, and converted into changes in metabolism, gene expression, secretion, motility, differentiation, immune activation, growth, apoptosis, or survival. Signaling is therefore one of the main ways life turns environmental and internal conditions into coordinated biological behavior.
Cell signaling also makes clear why biology cannot be understood through structure alone. A cell may contain DNA, proteins, membranes, and organelles, but its ability to live depends on regulation through time. The cell must interpret signals, prioritize responses, and coordinate change. Signaling is therefore one of the mechanisms through which living order becomes dynamic.
Why living systems must communicate
Living systems must communicate because life is conditional. Cells do not exist in fixed, unchanging environments, and multicellular organisms are not collections of independent units. Biological systems are dynamic and relational: they must match internal activity to external conditions and coordinate activity across components if they are to remain viable. Communication is therefore not an optional complexity added late to life. It is one of the central conditions of organized biological existence.
In unicellular organisms, signaling may regulate nutrient sensing, chemotaxis, stress response, biofilm formation, quorum sensing, sporulation, host interaction, and cooperative behavior. In multicellular organisms, signaling coordinates development, endocrine function, neural activity, immunity, wound repair, tissue growth, vascular regulation, metabolism, and homeostasis. At ecological scales, signaling-related behavior helps shape symbiosis, defense, reproduction, microbial cooperation, plant-microbe interaction, predator-prey response, and responses to disturbance.
Biology therefore treats signaling as a general principle rather than a niche topic. To understand signaling is to understand how life maintains coherence under conditions of uncertainty and change. A living system must not only contain molecules and cells. It must coordinate them.
This also explains why signaling appears across so many biological fields. Cell biologists study receptors and transduction. Physiologists study hormones, nerves, and homeostatic regulation. Immunologists study antigen recognition, cytokines, and inflammatory response. Ecologists study communication among organisms and response to environmental cues. Biotechnology researchers design assays, biosensors, and engineered response circuits. These fields differ in scale, but they share the same underlying problem: how information becomes biological action.
Signaling molecules and their receptors
Signaling molecules act by binding to receptors expressed by target cells. Some receptors are located on the cell surface, where they detect extracellular molecules that cannot readily cross the membrane. Others are intracellular receptors, which respond to small hydrophobic ligands capable of diffusing into the cytosol or nucleus. This receptor principle is foundational because it explains how signaling remains selective. A signaling molecule does not affect every nearby cell in the same way. It affects cells that express a compatible receptor and possess the downstream machinery needed to interpret the signal.
This selectivity is one reason the same signaling molecule can have different effects in different tissues, developmental states, immune contexts, or species interactions. A hormone, cytokine, neurotransmitter, or microbial signal is not meaningful by itself. Its meaning depends on receptor expression, pathway state, cellular history, concentration, duration, localization, and cross-talk with other signals.
Signaling molecules also operate across different spatial scales. Endocrine signals travel long distances through circulation. Paracrine signals act locally. Synaptic signaling is highly targeted. Autocrine signaling feeds back on the originating cell. Juxtacrine or contact-dependent signaling requires direct cellular interaction. Microbial quorum signals accumulate in shared environments and become meaningful when they cross concentration thresholds. Plant signaling molecules may travel through tissues or act locally during stress and development.
Receptors therefore function as biological interpreters. They do not merely bind molecules. They help determine whether a molecule becomes a signal and what kind of cellular response follows.
This interpretation depends on both specificity and context. A receptor can be present but inactive, desensitized, internalized, mutated, blocked, overexpressed, or embedded within a pathway state that changes its meaning. Signaling biology therefore treats receptor-ligand binding as the beginning of interpretation, not the entire response.
Major receptor types and signal transduction
Cell signaling depends on multiple receptor classes, but several broad categories are especially central. G protein-coupled receptors, or GPCRs, mediate responses to many hormones, neurotransmitters, sensory cues, chemokines, odorants, and local mediators through intermediary G proteins and downstream signaling systems. GPCR systems are important because they can translate ligand binding into changes in second messengers, ion channels, kinase pathways, and transcriptional states.
Enzyme-linked receptors, including receptor tyrosine kinases, act through ligand-triggered conformational changes and phosphorylation-dependent signaling cascades. These pathways are central to growth, differentiation, metabolism, survival, and development. Because phosphorylation can rapidly alter protein activity, localization, and interaction, kinase-mediated signaling provides one of the major mechanisms through which cells convert extracellular information into intracellular regulation.
Ion-channel-linked receptors convert signals into changes in membrane potential or ion flux. Intracellular receptors respond to ligands such as steroid hormones and other molecules capable of crossing membranes. Pattern-recognition receptors detect pathogen- or damage-associated molecular features. Mechanosensitive receptors and channels translate physical force into cellular response. Plant receptors, microbial sensors, and immune receptors further expand the signaling landscape.
What unites these systems is not identical structure but shared function: the conversion of signal detection into intracellular response. This conversion is what biologists mean by signal transduction. A receptor detects; a pathway translates; a cell responds.
Signal transduction is also where biological specificity becomes dynamic. A receptor may activate kinases, phosphatases, second messengers, transcription factors, ion channels, metabolic enzymes, or cytoskeletal components. The pathway outcome depends on which downstream elements are present, how they are localized, how long they remain active, and what other signals are present at the same time.
Second messengers, amplification, and intracellular response
Second messengers are intracellular signaling molecules that relay and amplify information from activated receptors to downstream effectors. Cyclic AMP, cyclic GMP, calcium ions, inositol trisphosphate, diacylglycerol, nitric oxide, and lipid-derived signaling intermediates are among the best-known examples. Their biological importance lies in amplification and integration: a relatively small external signal can generate a large intracellular response through cascades of second-messenger production, protein activation, and pathway branching.
Calcium signaling is especially important because transient increases in cytosolic Ca2+ can regulate secretion, contraction, transcription, metabolism, motility, fertilization, neuronal activity, and immune activation. Low resting cytosolic calcium is part of what makes calcium useful as a second messenger, because transient rises can carry informational meaning. Calcium signals may vary in amplitude, frequency, duration, localization, and subcellular distribution.
Second messengers therefore make signaling dynamic rather than merely binary. A pathway is not simply “on” or “off.” Signals can vary in strength, timing, oscillation, spatial confinement, and interaction with other pathways. Cells can interpret these differences in context-dependent ways.
This dynamic view is essential for understanding why signal duration matters. A short pulse may trigger reversible enzyme activation, while sustained activity may alter transcription, differentiation, proliferation, or cell fate. Signaling biology therefore depends not only on molecular identities but on temporal structure.
The same logic applies to biotechnology and experimental design. A single endpoint measurement may miss the most important signaling behavior if the biologically meaningful feature is peak amplitude, delay, duration, oscillation, recovery, or adaptation. Signaling is often a time-series problem, not merely a presence-or-absence problem.
Signaling networks, feedback, and biological control
Signaling pathways rarely operate as isolated linear chains. Instead, they form networks with feedback loops, feedforward structures, cross-pathway modulation, redundancy, pathway branching, scaffold proteins, compartmentalization, and context-sensitive regulation. This is why signaling outcomes depend not only on ligand-receptor binding, but also on cellular state, developmental history, metabolic condition, receptor abundance, localization, and competing pathway activity.
Feedback is especially important because it allows biological control. Negative feedback can limit pathway activation, restore homeostasis, prevent runaway response, and generate adaptation. Positive feedback can stabilize cell fate decisions, amplify immune activation, reinforce developmental transitions, or produce switch-like behavior. Feedforward motifs can filter noise, delay response, or anticipate downstream needs.
Cross-talk among GPCR, kinase, calcium, metabolic, immune, and transcriptional pathways further means that signaling often integrates rather than simply relays information. A cell usually receives many signals at once. The biological outcome depends on integration.
Modern cell signaling is therefore best understood as networked biological control rather than one-receptor-one-response simplicity. This is one reason signaling belongs naturally within systems biology. Signaling pathways are not only molecular mechanisms; they are dynamic control systems embedded in living organization.
This networked view also helps explain why signaling can be both robust and fragile. Redundant pathways may preserve function under perturbation, but feedback failure, chronic activation, receptor mutation, inflammatory amplification, or pathway rewiring can destabilize the system. Signaling control is therefore central to health, disease, ecological response, and biological engineering.
Cell signaling in physiology, development, and biological function
Cell signaling underlies physiology because coordinated biological function depends on regulated information transfer. Hormonal regulation, neural communication, immune surveillance, vascular response, secretion, contraction, growth, appetite control, circadian timing, tissue repair, reproduction, and stress response all require signaling systems. A physiological response is often best understood as a signaling event distributed across time, cell type, and tissue context.
Development also depends deeply on signaling. Cells must interpret positional information, respond to morphogens, communicate with neighbors, and stabilize fates through regulated pathways. A multicellular organism does not emerge merely because cells divide. It emerges because signaling pathways coordinate differentiation, polarity, timing, migration, growth, apoptosis, and form.
This makes signaling one of the main ways molecular biology becomes organismal biology. A receptor event at the membrane can alter gene expression; a morphogen gradient can help pattern tissue; an immune cytokine can reorganize cellular behavior; a hormone can coordinate distant organs; a neurotransmitter can shape behavior. Signaling is the connective architecture through which biological levels communicate.
In this sense, signaling is not one biological process among many. It is a principle of coordination across living systems.
This principle also explains why signaling is central to the study of biological failure. Developmental disorder, immune dysregulation, endocrine disruption, cancer, neurodegeneration, metabolic disease, and inflammatory pathology often involve failures of signal detection, interpretation, timing, or termination. A living system must not only respond. It must respond appropriately.
Ecology, communication, and sustainability-adjacent biology
Cell signaling is equally important in ecology because organisms do not respond to environments passively. They detect cues, assess resource conditions, regulate stress pathways, coordinate reproduction, alter behavior, and interact with other organisms through signaling processes. Ecological response is therefore partly a signaling problem expressed at larger scales.
This matters for sustainability-adjacent biology because environmental disruption becomes biologically real through signaling and response pathways. Heat stress, salinity change, drought, toxins, pathogens, nutrient limitation, hypoxia, acidification, and pollution all act partly by altering signaling states within cells and organisms. Resilience depends not only on external conditions, but on whether biological systems can detect change, interpret it accurately, and respond effectively.
Ecology thus benefits from a signaling perspective because it connects environmental pressure to physiological and molecular response. It shows how organisms and communities remain adaptive, vulnerable, or unstable under changing conditions. A changing environment does not simply surround life. It enters biology through detection, signaling, response, and failure.
This is especially important for conservation biology and restoration ecology. A population may decline not only because individuals die directly, but because environmental signals disrupt reproduction, migration, symbiosis, immunity, stress tolerance, or development. Signaling provides one of the mechanisms through which environmental change becomes biological consequence.
Marine, freshwater, soil, plant, and microbial relevance
Marine biology makes the importance of signaling especially visible. Marine organisms respond to temperature, salinity, light, oxygen availability, acidification, nutrient limitation, toxins, and symbiotic partners through signaling pathways that regulate metabolism, growth, stress tolerance, development, reproduction, and behavior. Coral bleaching, plankton response to nutrient regimes, microbial loop behavior, osmoregulatory adaptation, larval settlement, and host-symbiont coordination all depend partly on signaling-mediated response.
Freshwater biology presents similar signaling challenges under eutrophication, pollution, hypoxia, seasonal turnover, invasive species pressure, and hydrologic disturbance. Aquatic organisms must detect chemical cues, oxygen shifts, thermal stress, pathogens, and habitat signals in environments where chemical communication can be rapidly diluted, transformed, or disrupted.
Soil biology is equally signaling-rich. Roots, microbes, fungi, nematodes, and plant-associated communities coordinate nutrient exchange, defense, symbiosis, and decomposition through chemical communication. Mycorrhizal association, nitrogen-fixing symbiosis, root exudate signaling, microbial competition, and decomposition dynamics all involve signal production and interpretation.
Plant signaling more broadly links hormones, stress response, root architecture, stomatal regulation, defense, flowering, growth, and developmental plasticity under changing environmental conditions. Microbiology highlights perhaps the widest range of signaling strategies. Quorum sensing, biofilm formation, stress response, chemotaxis, host association, virulence regulation, and cooperative behavior all depend on signal production and interpretation. Microbial signaling is therefore central not only to molecular and cellular biology, but also to ecosystem function, agroecology, and biogeochemical cycling.
These examples show that signaling is not only a laboratory topic. It is embedded in the way organisms sense and respond to ecological worlds. Marine, freshwater, soil, plant, and microbial systems all demonstrate that biological communication is part of environmental adaptation.
Medical, biomedical, and disease ecology relevance
Cell signaling is foundational to medicine and biomedicine because disease often involves signaling failure, signaling overactivation, receptor alteration, pathway misregulation, feedback breakdown, or inappropriate cellular response. Cancer, inflammation, endocrine disorder, immune dysfunction, neurological disease, cardiovascular pathology, metabolic disease, fibrosis, and developmental disorders all depend heavily on signaling biology. GPCRs, receptor tyrosine kinases, cytokine receptors, nuclear receptors, ion channels, and intracellular signaling enzymes are especially important therapeutic and pharmacological targets.
Immunology is especially signaling-intensive. T cell activation, innate sensing, cytokine response, chemotaxis, antigen recognition, inflammatory resolution, and memory formation all depend on signaling cascades, second messengers, phosphorylation networks, calcium dynamics, and transcriptional reprogramming. A failure of immune signaling may produce immunodeficiency, autoimmunity, chronic inflammation, or ineffective pathogen clearance.
Disease ecology extends this further. Hosts, pathogens, vectors, and microbial communities all communicate chemically and respond through signaling systems. Infection, virulence, immune evasion, quorum-regulated pathogenicity, host manipulation, microbiome shifts, and ecological transmission are therefore not only population phenomena. They are also signaling phenomena expressed across scales.
This is why signaling is central to biomedical science and ecological health alike. It helps explain both the molecular basis of disease and the environmental contexts in which biological response succeeds or fails.
For medical and biomedical readers, signaling also provides a bridge between molecular mechanism and clinical outcome. A receptor mutation, cytokine storm, kinase activation, hormonal imbalance, or defective feedback loop can become a tissue-level disease, an organismal crisis, or a population-level health burden. Signaling connects mechanism to consequence.
Biotechnology, bioinformatics, and computational relevance
Biotechnology extends signaling biology into applied systems of intervention, assay design, engineering, and analysis. Reporter assays, receptor screening, kinase profiling, biosensors, synthetic signaling circuits, high-content imaging, calcium imaging, phosphoproteomics, pathway perturbation experiments, and drug discovery all depend on the ability to measure and manipulate signaling pathways in reproducible ways. In biotechnology, signaling becomes not only something to explain, but something to detect, perturb, engineer, and optimize.
Bioinformatics and computational biology are especially important because signaling networks are dynamic, branching, and difficult to interpret intuitively once they become large. Time-series data, pathway activation profiles, phosphoproteomic datasets, calcium traces, receptor-response curves, ligand panels, single-cell measurements, and gene-expression consequences often require computational analysis for visualization, modeling, clustering, and inference.
Synthetic biology strengthens this further by treating signaling modules as components that can be rewired, coupled, or redesigned. Engineered cells can be designed to sense disease states, environmental toxins, metabolites, or microbial signals and respond with programmed outputs. This makes signaling one of the clearest places where biological understanding becomes operational design.
Signaling therefore sits at a major intersection of experimental biology, computation, medicine, environmental sensing, and biological engineering.
This intersection also raises standards for reproducibility. A signaling experiment should document ligand identity, concentration, exposure time, receptor context, cell type, assay conditions, normalization method, time resolution, controls, and computational transformations. Without provenance, signaling data become difficult to interpret or compare.
Quantitative cell signaling: mathematics, R, and Python
Modern signaling biology is not only mechanistic. It is also quantitative. Receptor occupancy, second-messenger dynamics, pathway activation, feedback strength, signal duration, response thresholds, noise, oscillation, and pathway cross-talk can all be modeled mathematically and analyzed statistically. This does not replace biological understanding. It makes signaling behavior more explicit, testable, and comparable.
A simple receptor-occupancy relation can be written as:
Interpretation: Receptor occupancy links ligand concentration to the fraction of receptors engaged by ligand.
where \(\theta\) is the fraction of occupied receptors, \([L]\) is ligand concentration, and \(K_d\) is the dissociation constant. This is useful because it links signal concentration to receptor engagement and helps explain thresholds, sensitivity, and saturation.
Many signaling systems are better approximated by a Hill-type response curve:
Interpretation: A Hill response captures cooperative, switch-like, or ultrasensitive signaling behavior.
where \(R(L)\) is normalized response, \(L\) is ligand concentration, \(K\) is the half-response level, and \(n\) is the Hill coefficient. This is useful because it captures switch-like behavior, cooperative binding, or ultrasensitive pathway response more realistically than a simple linear model.
A simple exponential-decay model for a transient intracellular messenger is:
Interpretation: Messenger decay models the disappearance or clearance of a transient intracellular signal.
where \(M(t)\) is messenger abundance at time \(t\), \(M_0\) is initial abundance, and \(k\) is the decay constant. The corresponding half-life is:
Interpretation: The half-life is the time required for the signal to decline to half its initial value.
This is useful for thinking about transient signaling pulses, clearance, and response duration.
A simple input-output signaling model can be written as:
Interpretation: Pathway activity rises with input strength and declines through decay or deactivation.
where \(S\) is pathway activity, \(I(t)\) is a time-varying input pulse, \(\alpha\) is activation strength, and \(\beta\) is decay or deactivation rate. This is useful because many signaling pathways are shaped not only by signal presence, but by pulse duration, timing, and dissipation.
A minimal signaling-feedback system can be represented as:
Interpretation: Signaling activity can induce feedback inhibition that dampens or adapts the response over time.
where \(S\) is signaling activity and \(F\) is a feedback inhibitor. This is useful because many real signaling systems generate their own dampening factors, producing adaptation, homeostasis, and transient responses instead of perpetual activation.
A simple quorum signal can be represented as accumulation with degradation:
Interpretation: Quorum signal concentration increases with population density and decreases through degradation or dilution.
where \(Q\) is signal concentration, \(N\) is population density, \(a\) is signal production per cell, and \(d\) is degradation or dilution. A response may occur when:
Interpretation: Quorum-regulated behavior begins when the accumulated signal crosses an activation threshold.
where \(Q_c\) is the activation threshold. This is useful because microbial communication often depends on signal accumulation relative to density, diffusion, and decay.
Variables, units, and signaling interpretation
Quantitative signaling biology depends on variables that connect molecular concentration, receptor engagement, pathway activation, decay, feedback, and threshold response. The table below summarizes several central quantities.
| Symbol or Term | Meaning | Typical Unit or Scale | Signaling Interpretation |
|---|---|---|---|
| \([L]\) or \(L\) | Ligand concentration | molarity, mass per volume, arbitrary assay units, or relative concentration | External or internal signal level available for receptor binding or pathway activation |
| \(K_d\) | Dissociation constant | same concentration unit as ligand | Ligand concentration at which half of receptors are occupied in a simple binding model |
| \(\theta\) | Receptor occupancy | fraction from 0 to 1 | Share of receptors bound by ligand |
| \(R(L)\) | Normalized signaling response | fraction or normalized response | Downstream pathway activation as a function of ligand level |
| \(K\) | Half-response level | same concentration unit as ligand | Ligand level associated with half-maximal pathway response |
| \(n\) | Hill coefficient | dimensionless | Controls steepness or cooperativity of signaling response |
| \(M(t)\) | Messenger abundance at time \(t\) | concentration, fluorescence intensity, or arbitrary units | Level of a transient intracellular messenger over time |
| \(M_0\) | Initial messenger abundance | same as \(M(t)\) | Starting signal amplitude before decay |
| \(k\) | Decay constant | per unit time | Rate at which a signaling molecule or activity decays |
| \(t_{1/2}\) | Half-life | time | Time required for a signaling quantity to decline by half |
| \(S\) | Pathway activity | normalized activity, fluorescence, phosphorylation score, or arbitrary units | Dynamic state of a signaling pathway |
| \(I(t)\) | Time-varying input | concentration, binary pulse, or normalized input | Signal stimulus applied over time |
| \(F\) | Feedback inhibitor | normalized level or concentration | Inhibitory component induced by signaling activity |
| \(Q\) | Quorum signal concentration | concentration or normalized signal level | Accumulated microbial communication signal |
| \(Q_c\) | Quorum activation threshold | same as \(Q\) | Signal level at which quorum-regulated response begins |
The table illustrates why signaling data require careful interpretation. The same measured response may represent receptor binding, second-messenger abundance, enzyme activity, transcriptional output, fluorescence intensity, or population-level communication. Mathematical variables become biologically meaningful only when linked to measurement context.
Worked example: signaling half-life and receptor occupancy
Suppose a second messenger starts at \(M_0=100\) arbitrary units and declines to 25 units after 4 minutes. Under the decay model:
Interpretation: The messenger level declines exponentially as the signal is degraded, cleared, or deactivated.
Substituting the values:
Interpretation: The observed signal trace can be used to estimate the decay constant.
Dividing both sides by 100:
Interpretation: The messenger has declined to 25 percent of its initial level after 4 minutes.
Taking the natural logarithm:
Interpretation: The decay constant can be estimated by linearizing the exponential decay relation.
Solving gives:
Interpretation: The estimated decay constant is approximately 0.3466 per minute.
The half-life is:
Interpretation: The messenger signal halves approximately every 2 minutes under these conditions.
This is useful because it converts a signaling trace into an interpretable dynamic parameter rather than a purely descriptive graph.
Receptor occupancy can be interpreted similarly. Suppose ligand concentration is \([L]=3\) and the receptor dissociation constant is \(K_d=1.5\). Then:
Interpretation: About 66.7 percent of receptors are occupied at this ligand concentration under the simple binding model.
This gives a simple way to interpret signaling sensitivity and relative receptor engagement under changing ligand conditions.
R and Python workflows
Computational modeling helps make signaling biology explicit because signaling is dynamic, threshold-sensitive, and often nonlinear. A receptor-occupancy model can show saturation. A Hill curve can show cooperative response. A decay model can estimate signaling half-life. A pulse-driven system can show transient activation. A feedback model can show adaptation or self-limitation. A quorum-sensing model can show how population density becomes collective behavior through signal accumulation.
The selected examples below focus on compact, reusable workflows that can be adapted for receptor assays, reporter systems, calcium traces, phosphoproteomic time series, microbial quorum sensing, and synthetic signaling circuits. The GitHub repository extends this into broader computational scaffolding with SQL provenance structures, validation notes, data files, notebooks, and multi-language implementations.
The goal is not to make signaling abstract. The goal is to make signaling evidence traceable. A signaling claim becomes stronger when the ligand, receptor, dose, time, model, parameters, assumptions, and output can be inspected together.
R workflow: occupancy, decay, feedback, and quorum thresholds
R is useful for signaling analysis because it supports statistical fitting, time-series summaries, threshold detection, and reproducible tabular workflows. The following workflow compares receptor occupancy with cooperative response, estimates signaling half-life, simulates negative feedback, and models quorum-sensing threshold behavior.
# Cell Signaling Quantitative Workflow
#
# This workflow demonstrates four signaling-analysis tasks:
#
# 1. Compare receptor occupancy with a cooperative Hill response.
# 2. Estimate second-messenger decay and half-life.
# 3. Simulate pulse-driven signaling with negative feedback.
# 4. Simulate quorum-sensing signal accumulation and threshold crossing.
#
# These examples can be adapted for receptor assays, reporter assays,
# calcium/cAMP time series, kinase activation traces, microbial quorum sensing,
# biosensor response curves, or synthetic signaling circuits.
library(tibble)
library(dplyr)
# ------------------------------------------------------------
# 1. Receptor occupancy and Hill response
# ------------------------------------------------------------
response_df <- tibble(
ligand = seq(0.01, 10, length.out = 300)
) %>%
mutate(
Kd = 1.5,
K = 2.0,
hill_n = 3.0,
occupancy = ligand / (Kd + ligand),
hill_response = ligand^hill_n / (K^hill_n + ligand^hill_n)
)
threshold_row <- response_df %>%
slice(which.min(abs(hill_response - 0.5)))
# ------------------------------------------------------------
# 2. Fit an exponential messenger-decay model
# ------------------------------------------------------------
decay_df <- tibble(
time_min = c(0, 1, 2, 3, 4),
signal = c(100, 70, 50, 35, 25)
)
decay_model <- lm(log(signal) ~ time_min, data = decay_df)
decay_summary <- tibble(
k_est = -coef(decay_model)[["time_min"]],
m0_est = exp(coef(decay_model)[["(Intercept)"]]),
half_life_min = log(2) / k_est,
r_squared_log_space = summary(decay_model)$r.squared
)
decay_fit <- decay_df %>%
mutate(
predicted_signal = exp(predict(decay_model)),
residual_log_scale = resid(decay_model)
)
# ------------------------------------------------------------
# 3. Pulse-driven signaling with negative feedback
# ------------------------------------------------------------
times <- seq(0, 20, by = 0.1)
feedback_df <- tibble(
time = times,
input_signal = ifelse(times >= 2 & times <= 8, 1, 0),
pathway_activity = 0,
feedback_inhibitor = 0
)
alpha <- 3.0
beta <- 0.6
gamma <- 0.15
delta <- 0.4
epsilon <- 0.3
for (i in 2:nrow(feedback_df)) {
dt <- feedback_df$time[i] - feedback_df$time[i - 1]
dS <- alpha * feedback_df$input_signal[i - 1] -
beta * feedback_df$pathway_activity[i - 1] -
gamma * feedback_df$feedback_inhibitor[i - 1] *
feedback_df$pathway_activity[i - 1]
dF <- delta * feedback_df$pathway_activity[i - 1] -
epsilon * feedback_df$feedback_inhibitor[i - 1]
feedback_df$pathway_activity[i] <-
max(feedback_df$pathway_activity[i - 1] + dS * dt, 0)
feedback_df$feedback_inhibitor[i] <-
max(feedback_df$feedback_inhibitor[i - 1] + dF * dt, 0)
}
peak_feedback_row <- feedback_df %>%
slice(which.max(pathway_activity))
# ------------------------------------------------------------
# 4. Quorum-sensing threshold simulation
# ------------------------------------------------------------
quorum_df <- tibble(
time = seq(0, 24, by = 0.1)
) %>%
mutate(
population_density = pmin(1e5 * exp(0.25 * time), 1e9),
quorum_signal = 0,
response_active = FALSE
)
signal_production_per_cell <- 1e-9
signal_decay <- 0.35
activation_threshold <- 1.0
for (i in 2:nrow(quorum_df)) {
dt <- quorum_df$time[i] - quorum_df$time[i - 1]
dQ <- signal_production_per_cell * quorum_df$population_density[i - 1] -
signal_decay * quorum_df$quorum_signal[i - 1]
quorum_df$quorum_signal[i] <-
max(quorum_df$quorum_signal[i - 1] + dQ * dt, 0)
}
quorum_df <- quorum_df %>%
mutate(response_active = quorum_signal >= activation_threshold)
first_quorum_row <- quorum_df %>%
filter(response_active) %>%
slice_head(n = 1)
print(head(response_df, 12))
print(tail(response_df, 12))
print(threshold_row)
print(round(decay_summary, 4))
print(round(decay_fit, 4))
print(head(feedback_df, 15))
print(tail(feedback_df, 15))
print(round(peak_feedback_row, 4))
print(head(quorum_df, 10))
print(round(first_quorum_row, 4))This workflow is useful because it moves from receptor engagement to dynamic pathway behavior. It shows how signaling can saturate, decay, adapt, and cross thresholds rather than behaving as a static binary switch.
Python workflow: receptor response, half-life, feedback, and quorum sensing
Python is useful for signaling biology because it supports simulation, parameter sweeps, pipeline design, data validation, and reproducible modeling. The following workflow computes receptor occupancy and Hill response, estimates signaling half-life, simulates pulse-driven signaling with feedback, and models quorum-sensing threshold crossing.
"""
Cell Signaling Computational Workflow
This workflow demonstrates four common quantitative signaling tasks:
1. Compare receptor occupancy with a cooperative Hill response.
2. Estimate signaling decay and half-life from a time course.
3. Simulate pulse-driven pathway activity with negative feedback.
4. Simulate quorum-sensing signal accumulation and threshold crossing.
The examples are compact, but the same structures can be extended to
receptor assays, reporter systems, calcium/cAMP time series,
phosphorylation dynamics, microbial quorum sensing, biosensors,
and synthetic signaling circuits.
"""
from __future__ import annotations
import numpy as np
import pandas as pd
def receptor_response(
kd: float = 1.5,
k_half: float = 2.0,
hill_n: float = 3.0,
max_ligand: float = 10.0,
n_points: int = 300,
) -> pd.DataFrame:
"""
Calculate receptor occupancy and a cooperative Hill response.
"""
ligand = np.linspace(0.01, max_ligand, n_points)
occupancy = ligand / (kd + ligand)
hill_response = ligand**hill_n / (k_half**hill_n + ligand**hill_n)
return pd.DataFrame(
{
"ligand": ligand,
"occupancy": occupancy,
"hill_response": hill_response,
}
)
def fit_signaling_half_life() -> tuple[pd.DataFrame, pd.DataFrame]:
"""
Estimate exponential signaling decay and half-life.
"""
time_min = np.array([0, 1, 2, 3, 4], dtype=float)
signal = np.array([100, 70, 50, 35, 25], dtype=float)
slope, intercept = np.polyfit(time_min, np.log(signal), 1)
k_est = -slope
m0_est = np.exp(intercept)
half_life = np.log(2.0) / k_est
predicted = np.exp(intercept + slope * time_min)
summary = pd.DataFrame(
{
"k_est": [k_est],
"m0_est": [m0_est],
"half_life_min": [half_life],
}
)
trace = pd.DataFrame(
{
"time_min": time_min,
"observed_signal": signal,
"predicted_signal": predicted,
}
)
return summary, trace
def simulate_feedback(
t_max: float = 20.0,
dt: float = 0.1,
alpha: float = 3.0,
beta: float = 0.6,
gamma: float = 0.15,
delta: float = 0.4,
epsilon: float = 0.3,
) -> pd.DataFrame:
"""
Simulate pulse-driven signaling with induced negative feedback.
"""
times = np.arange(0, t_max + dt, dt)
pathway_activity = np.zeros_like(times)
feedback_inhibitor = np.zeros_like(times)
input_signal = np.where((times >= 2.0) & (times <= 8.0), 1.0, 0.0)
for i in range(1, len(times)):
step = times[i] - times[i - 1]
d_pathway = (
alpha * input_signal[i - 1]
- beta * pathway_activity[i - 1]
- gamma * feedback_inhibitor[i - 1] * pathway_activity[i - 1]
)
d_feedback = delta * pathway_activity[i - 1] - epsilon * feedback_inhibitor[i - 1]
pathway_activity[i] = max(pathway_activity[i - 1] + d_pathway * step, 0.0)
feedback_inhibitor[i] = max(feedback_inhibitor[i - 1] + d_feedback * step, 0.0)
return pd.DataFrame(
{
"time": times,
"input_signal": input_signal,
"pathway_activity": pathway_activity,
"feedback_inhibitor": feedback_inhibitor,
}
)
def simulate_quorum_sensing(
t_max: float = 24.0,
dt: float = 0.1,
initial_density: float = 1e5,
population_growth_rate: float = 0.25,
max_density: float = 1e9,
signal_production_per_cell: float = 1e-9,
signal_decay: float = 0.35,
activation_threshold: float = 1.0,
) -> pd.DataFrame:
"""
Simulate quorum-sensing signal accumulation and threshold response.
"""
times = np.arange(0, t_max + dt, dt)
population_density = initial_density * np.exp(population_growth_rate * times)
population_density = np.minimum(population_density, max_density)
quorum_signal = np.zeros_like(times)
for i in range(1, len(times)):
step = times[i] - times[i - 1]
d_signal = (
signal_production_per_cell * population_density[i - 1]
- signal_decay * quorum_signal[i - 1]
)
quorum_signal[i] = max(quorum_signal[i - 1] + d_signal * step, 0.0)
return pd.DataFrame(
{
"time": times,
"population_density": population_density,
"quorum_signal": quorum_signal,
"response_active": quorum_signal >= activation_threshold,
}
)
def main() -> None:
"""
Run compact signaling workflows.
"""
response_df = receptor_response()
threshold_row = response_df.iloc[
(response_df["hill_response"] - 0.5).abs().argmin()
]
decay_summary, decay_trace = fit_signaling_half_life()
feedback_df = simulate_feedback()
quorum_df = simulate_quorum_sensing()
first_quorum_row = quorum_df.loc[quorum_df["response_active"]].head(1)
print("Receptor occupancy and Hill response:")
print(response_df.head(12).round(4).to_string(index=False))
print(response_df.tail(12).round(4).to_string(index=False))
print("\nApproximate half-response row:")
print(threshold_row.round(4).to_string())
print("\nSignaling half-life estimate:")
print(decay_summary.round(4).to_string(index=False))
print(decay_trace.round(4).to_string(index=False))
print("\nPulse-driven signaling with feedback:")
print(feedback_df.head(15).round(4).to_string(index=False))
print(feedback_df.tail(15).round(4).to_string(index=False))
print("\nPeak pathway activity:")
print(feedback_df.loc[feedback_df["pathway_activity"].idxmax()].round(4).to_string())
print("\nQuorum-sensing threshold simulation:")
print(quorum_df.head(10).round(4).to_string(index=False))
print("\nFirst threshold crossing:")
print(first_quorum_row.round(4).to_string(index=False))
if __name__ == "__main__":
main()This Python workflow is useful because it introduces the logic of adaptive signaling rather than treating pathways as static or purely linear chains. It also shows how receptor binding, response steepness, signal decay, feedback inhibition, and microbial threshold behavior can be modeled with transparent assumptions.
GitHub repository
The article body includes compact R and Python examples so the biological and scientific argument remains readable. The full repository expands those examples into a broader computational cell-signaling workflow, including receptor occupancy, Hill response curves, second-messenger decay, signaling half-life estimation, pulse-driven pathway dynamics, negative-feedback models, quorum-sensing threshold simulation, calcium/cAMP-style time-series scaffolds, pathway activation scoring, SQL provenance structures, reproducible data files, and full-stack scientific-computing examples across Python, R, Julia, Fortran, Rust, Go, C, C++, SQL, and notebooks.
Complete Code Repository
The full code distribution for this article, including selected article examples, expanded computational workflows, reproducible data structures, provenance documentation, and full-stack scientific-computing scaffolding, is available on GitHub.
Limits, scaling, and modern signaling thinking
Signaling biology is foundational, but it is not simple. The same receptor can produce different outcomes in different cell types. The same ligand can have different effects depending on dose, duration, receptor density, pathway state, cellular history, metabolic condition, or environmental context. A signaling cascade characterized cleanly in vitro may behave differently in tissues, microbiomes, soils, aquatic systems, or whole organisms. Context is not a complication added after the fact. It is part of what signaling means biologically.
This is why modern signaling thinking increasingly emphasizes networks, timescales, localization, stochasticity, feedback, cross-talk, and system context. Signaling is rarely a single linear path from ligand to response. It is often a distributed control system in which multiple inputs are integrated, weighted, transformed, amplified, dampened, and interpreted under constraints set by history, environment, and physiology.
Models and workflows are useful because they clarify assumptions, expose mechanisms, and make comparison possible. But a receptor-occupancy equation is not a complete signaling pathway, a Hill curve is not a full cellular response, and a feedback model is not a complete tissue or ecological system. Quantitative signaling biology is strongest when it supports biological interpretation rather than replacing it.
In that sense, signaling is a model case for modern biology itself: mechanistic, dynamic, multi-scale, environmentally embedded, computationally analyzable, and irreducible to a single level of explanation.
This caution matters in experimental and applied settings. A drug response curve may not predict tissue outcome. A pathway activation score may not capture cell fate. A biosensor may behave differently in environmental conditions than in controlled media. Signaling data are powerful, but they must remain connected to biological context.
Why this matters for scientific work
For working scientists, signaling matters because many biological problems are misread when communication pathways are treated as black boxes. A developmental problem may depend on how a morphogen gradient is interpreted rather than on the mere presence of developmental genes. A plant stress problem may depend on hormonal cross-talk and threshold sensitivity rather than on one isolated stress pathway. A disease problem may depend on feedback failure or persistent pathway activation rather than on gross structural damage alone. A microbial ecology problem may depend on quorum thresholds and signal accumulation rather than on abundance alone.
This means signaling should often be treated as explanatory infrastructure rather than as a narrow subsection of cell biology. Physiologists need it because coordinated function depends on regulated information transfer. Developmental biologists need it because cell fate and patterning are signaling-intensive. Ecologists need it because organisms and communities respond to change through communication pathways. Biomedical scientists need it because many diseases are disorders of signaling control. Computational biologists need it because signaling is one of the clearest dynamic network problems in modern biology.
The scientific importance of signaling lies partly in this breadth. It is one of the principal ways biology explains how living systems coordinate themselves under changing conditions.
Signaling also matters because it connects mechanism to intervention. If a pathway can be measured, perturbed, modeled, and validated, it can become a therapeutic target, biosensor design, environmental indicator, engineering module, or diagnostic signal. That makes signaling one of the most important bridges between biological understanding and applied biological power.
Conclusion
Cell signaling, communication, and biological coordination show that life depends not only on matter, energy, and genetic information, but also on regulated information flow. Cells and organisms must detect cues, interpret conditions, amplify signals, regulate pathways, and coordinate responses if they are to remain viable, adaptive, and organized.
To understand signaling is therefore to understand one of the deepest conditions of life: that biological systems are communicative systems. Signaling links receptors to physiology, second messengers to behavior, pathway networks to development, and cellular response to ecological and evolutionary consequence. That is why signaling remains central not only to cell biology and physiology, but also to ecology, marine and freshwater biology, soil biology, plant science, microbiology, disease ecology, medicine, and biotechnology.
Cell signaling is thus more than a topic within molecular biology. It is one of the principal ways biology explains how living systems coordinate themselves across scales. Its scientific importance lies in the fact that it connects detection to response, molecules to systems, and environmental change to biological meaning.
Related articles
- Biology
- Cell Theory and the Basic Unit of Life
- DNA, RNA, and the Molecular Logic of Life
- Molecular Biology and the Flow of Genetic Information
- Enzymes, Regulation, and Biochemical Pathways
- Metabolism, Energy, and Biological Function
- Epigenetics, Regulation, and Gene Expression
- Physiology and the Regulation of Living Systems
- Immunology and Biological Defense
- Microbiology and the Hidden Majority of Life
- Systems Biology and the Logic of Biological Integration
Further reading
- Alberts, B. et al. (2002) ‘Signaling molecules and their receptors’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9924/
- Alberts, B. et al. (2002) ‘Functions of cell-surface receptors’, in Molecular Biology of the Cell. 4th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9866/
- Alberts, B. et al. (2002) ‘Pathways of intracellular signal transduction’, in Molecular Biology of the Cell. 4th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9870/
- Alberts, B. et al. (2002) ‘Signaling through G-protein-linked cell-surface receptors’, in Molecular Biology of the Cell. 4th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26912/
- Alberts, B. et al. (2002) ‘Signaling through enzyme-linked cell-surface receptors’, in Molecular Biology of the Cell. 4th edn. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26822/
- NCBI Bookshelf (n.d.) Second Messengers. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10794/
- NCBI Bookshelf (n.d.) Ca2+ Signaling. Available at: https://www.ncbi.nlm.nih.gov/books/NBK27950/
- Khalil, B. et al. (2024) ‘Physiology, cellular receptors’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK554403/
- Grimes, A.C. et al. (2023) ‘Biochemistry, G protein coupled receptors’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK518966/
- Patra, C. et al. (2023) ‘Biochemistry, cAMP’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK535431/
- Pasmanter, N. et al. (2022) ‘Biochemistry, cyclic GMP’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK542234/
- Cooper, D. (2023) ‘Biochemistry, calcium channels’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK562198/
- Wintheiser, G.A. et al. (2022) ‘Physiology, tyrosine kinase receptors’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK538532/
References
- Alberts, B. et al. (2002) ‘Functions of cell-surface receptors’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9866/
- Alberts, B. et al. (2002) ‘Pathways of intracellular signal transduction’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9870/
- Alberts, B. et al. (2002) ‘Signaling molecules and their receptors’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK9924/
- Alberts, B. et al. (2002) ‘Signaling through enzyme-linked cell-surface receptors’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26822/
- Alberts, B. et al. (2002) ‘Signaling through G-protein-linked cell-surface receptors’, in Molecular Biology of the Cell. 4th edn. New York: Garland Science. Available at: https://www.ncbi.nlm.nih.gov/books/NBK26912/
- Cooper, D. (2023) ‘Biochemistry, calcium channels’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK562198/
- Grimes, A.C. et al. (2023) ‘Biochemistry, G protein coupled receptors’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK518966/
- Khalil, B. et al. (2024) ‘Physiology, cellular receptors’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK554403/
- NCBI Bookshelf (n.d.) Ca2+ Signaling. Available at: https://www.ncbi.nlm.nih.gov/books/NBK27950/
- NCBI Bookshelf (n.d.) Second Messengers. Available at: https://www.ncbi.nlm.nih.gov/books/NBK10794/
- Pasmanter, N. et al. (2022) ‘Biochemistry, cyclic GMP’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK542234/
- Patra, C. et al. (2023) ‘Biochemistry, cAMP’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK535431/
- Wintheiser, G.A. et al. (2022) ‘Physiology, tyrosine kinase receptors’, in StatPearls. Treasure Island, FL: StatPearls Publishing. Available at: https://www.ncbi.nlm.nih.gov/books/NBK538532/