
Last Updated May 20, 2026
Sensory memory is the earliest stage of information processing, in which raw sensory input is briefly retained after stimulation has ended. These short-lived traces allow perceptual systems to stabilize incoming information long enough for attentional mechanisms to select what matters for further processing. Without sensory memory, experience would be far more fragmented, because sensory signals would disappear before the cognitive system could integrate them into coherent perception.
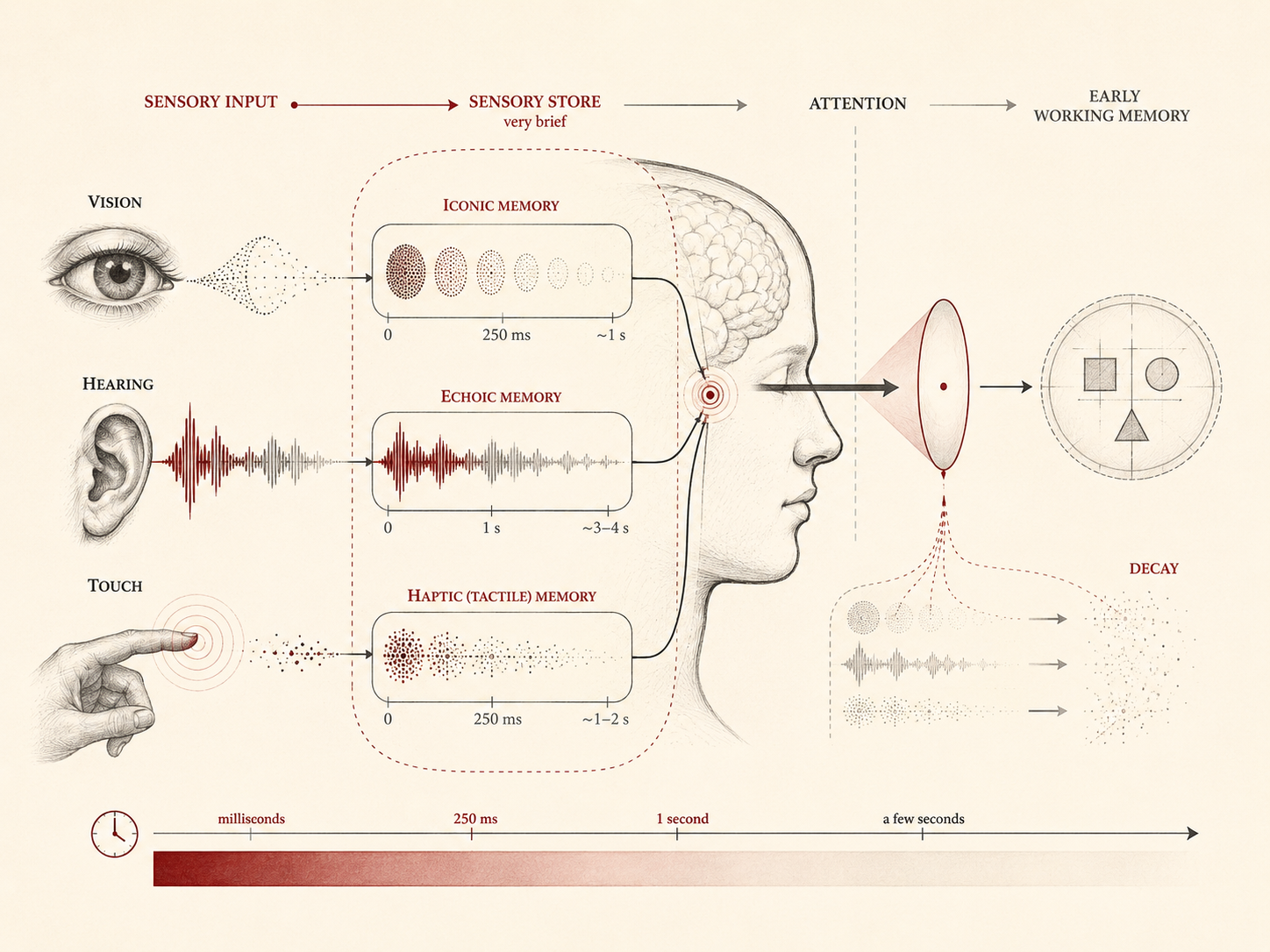
In cognitive psychology, sensory memory occupies the first stage of the broader processing architecture that transforms environmental stimulation into perception, attention, working memory, thought, and action. Incoming sensory signals are captured momentarily by modality-specific sensory registers associated with vision, hearing, touch, and related systems. These registers retain highly detailed but extremely brief representations of sensory input, usually for intervals measured in fractions of a second or, in some modalities, a few seconds.
Sensory memory therefore functions as a transitional buffer between sensation and higher cognition. Information retained in sensory memory may fade rapidly, or it may be selected through attention and stabilized for further processing in working memory. This filtering process is essential because the sensory environment contains far more information than can be processed consciously at once.
Main Library
Publications
Article Map
Cognitive Psychology
Related Topic
Behavioral Economics
Related Topic
Artificial Intelligence Systems
Related Topic
Data Systems & Analytics
The importance of sensory memory is easy to underestimate because it is so brief. Yet this brevity is precisely what makes it powerful. Sensory memory gives the cognitive system a short window in which the world remains available after the physical signal has changed or disappeared. It is not durable memory in the ordinary sense, but it is one of the first mechanisms through which stimulation becomes experience.
What is sensory memory?
Sensory memory refers to the earliest retention of sensory information after a stimulus has been presented. Unlike working memory or long-term memory, it does not maintain information in an actively manipulable or conceptually organized form. Instead, it preserves a fleeting sensory trace that allows the system to register and stabilize what has just occurred.
This brief retention matters because sensory input unfolds across time. Vision is interrupted by eye movements, audition is sequential, and touch often depends on changing contact across the body. Sensory memory gives cognition a short temporal bridge across these disruptions, making it possible for perception to feel continuous rather than disjointed.
Sensory memory is not the same as conscious recollection. A visual trace may be available for selection without being fully reportable. An auditory trace may persist long enough to support speech comprehension without becoming an explicit memory. A tactile trace may help integrate touch across movement without becoming a verbally accessible representation.
This distinction is central. Sensory memory is a preconceptual and often preconscious stage of retention. It preserves the recent sensory world long enough for attention and perceptual organization to operate. Some information is selected and stabilized. Much of it fades.
In this sense, sensory memory is not simply an incidental first stage. It is one of the conditions that make coherent perception possible at all.
The structure of sensory memory
Sensory memory is not a single undifferentiated system. It is better understood as a family of modality-specific registers associated with different sensory systems. Each register preserves a transient representation of incoming stimulation long enough for perceptual organization and attentional selection to occur.
The most widely discussed forms are:
- Iconic memory — sensory persistence for visual input.
- Echoic memory — sensory persistence for auditory input.
- Haptic memory — sensory persistence for tactile input.
These systems differ in duration, format, and function. Iconic memory is typically extremely brief but high in detail. Echoic memory tends to last longer, which is useful for speech and sound sequences. Haptic memory is less often discussed in introductory treatments, but it is important for the persistence of tactile input and the organization of touch-based experience.
Because sensory memory is modality-specific, it should not be treated as a single central storage box. Vision, audition, and touch each present different temporal problems. Visual input can be spatially rich but rapidly overwritten. Auditory input is sequential and must persist long enough to support temporal integration. Tactile input is distributed across the body and often depends on active exploration.
What these systems share is a common functional role: they briefly preserve recent sensory information so that the cognitive system can select, organize, and route it into later processing.
Iconic memory and the visual register
Research on iconic memory was transformed by George Sperling’s classic experiments on briefly presented letter arrays. Participants were often unable to report all the letters they had seen, but partial-report cueing showed that much more visual information had been momentarily available than whole-report performance suggested.
In a typical partial-report paradigm, participants see a brief array of letters and then receive a cue indicating which row to report. When the cue follows the display very quickly, performance is better than whole report would predict. As the cue delay increases, the advantage fades. This pattern suggests that a rich visual trace exists briefly after stimulus offset but decays rapidly before all of it can be reported.
Iconic memory therefore acts as a temporary visual store that preserves sensory input long enough for attentional processes to select what should be stabilized for further use. It is high in detail but short in duration. The richness of the trace does not mean that all of it becomes conscious or reportable. Rather, it shows that early visual availability exceeds later report capacity.
Later work complicated the earliest picture by distinguishing among different forms of visual persistence. Coltheart’s influential review separated iconic memory from visible persistence and emphasized that visual persistence, neural persistence, and informational persistence should not be treated as the same phenomenon. This matters because an afterimage, a briefly available visual representation, and a reportable memory trace may involve different levels of processing.
Iconic memory remains central because it reveals a basic architecture of cognition: early sensory systems can preserve more information than later systems can retain, report, or use.
Echoic memory and auditory persistence
Echoic memory refers to the auditory sensory register that briefly preserves sound information after the stimulus has ended. This persistence is essential for speech, music, and environmental sound processing because auditory input arrives sequentially rather than all at once.
When listening to speech, the brain must integrate successive phonemes, syllables, words, rhythms, and pauses into meaningful phrases and sentences. Echoic memory allows recently heard sounds to remain available long enough for that integration to occur. This longer temporal window is one reason auditory experience can preserve meaningful sequences even when attention shifts slightly or incoming sound is distributed over time.
Darwin, Turvey, and Crowder developed an auditory analogue of Sperling’s partial-report procedure. Their work showed that partial report could exceed whole report when auditory material was presented across spatial channels and cued soon after presentation. This supported the idea of brief auditory sensory storage, especially when spatial location served as the reporting cue.
Compared with iconic memory, echoic memory is often described as longer-lasting. This difference makes functional sense. Visual arrays can present spatial structure simultaneously, whereas auditory information must be integrated across time. Speech and sound perception require the system to retain recent auditory input long enough to interpret temporal patterns.
Echoic memory therefore helps explain how auditory cognition bridges the gap between transient sound waves and stable comprehension. Without such persistence, speech and music would be far harder to organize into meaningful sequences.
Haptic memory and tactile persistence
Haptic memory refers to the brief retention of tactile information. Although it receives less attention than iconic and echoic memory in many introductory accounts, it is important for understanding the continuity of touch, manual exploration, bodily orientation, and interaction with objects.
Tactile perception often depends on sequences of contact distributed across movement and time. A person exploring a surface with the hand receives changing information about pressure, texture, contour, vibration, temperature, and resistance. Haptic sensory memory helps preserve recent traces long enough for these changing signals to be integrated into a more coherent representation.
Haptic memory is especially relevant when touch must support recognition, navigation, or action. Manual object recognition, Braille reading, tool use, and tactile warning systems all depend on the integration of information across contact and motion. In these contexts, sensory persistence is not a theoretical curiosity. It is part of how bodily interaction becomes perceptual knowledge.
Research on haptic memory remains less developed than research on iconic and echoic memory, but contemporary work has begun to examine tactile persistence using change detection, somatosensory responses, and other experimental paradigms. This growing literature shows that sensory memory should be understood across the full range of sensory systems, not only vision and audition.
Haptic memory also has important implications for accessibility. Interfaces, warning systems, assistive technologies, and tactile displays should not assume that tactile information is processed identically to visual or auditory information. Touch has its own timing, spatial organization, and attentional constraints.
Formalizing sensory memory: decay, selection, and buffering
Sensory memory can be described formally as a rapidly decaying trace that preserves recent input for a short interval. Let \(s(t)\) represent the strength of a sensory trace at time \(t\) after stimulus offset. A simple exponential decay model is:
s(t)=s_0e^{-\lambda t}
\]
Interpretation: Trace strength \(s(t)\) begins at initial strength \(s_0\) and decays over time according to the decay-rate parameter \(\lambda\).
Because sensory registers differ across modalities, a modality-specific version can be written as:
s_m(t)=s_{0m}e^{-\lambda_m t}
\]
Interpretation: Each modality \(m\), such as vision, audition, or touch, may have its own initial trace strength and decay rate.
Selection from sensory memory can also be represented probabilistically. If a sensory trace \(i\) has strength \(s_i\), salience \(v_i\), and attentional priority \(a_i\), then the probability that it is selected for further processing can be expressed as:
Pr(i)=\frac{\exp(\beta_1s_i+\beta_2v_i+\beta_3a_i)}{\sum_{j=1}^{n}\exp(\beta_1s_j+\beta_2v_j+\beta_3a_j)}
\]
Interpretation: The probability of selecting trace \(i\) depends on trace strength, bottom-up salience, and top-down attentional priority relative to competing traces.
One can also represent the transition from sensory memory into later systems as a threshold process. If a trace exceeds a selection threshold \(\theta\), it is forwarded to later processing:
T_i=
\begin{cases}
1, & \text{if } s_i+a_i\geq\theta\\
0, & \text{otherwise}
\end{cases}
\]
Interpretation: \(T_i=1\) indicates that a trace has been selected or stabilized for a later stage such as working memory.
Partial-report advantage can be expressed as:
A_{PR}=R_{\text{partial}}-R_{\text{whole}}
\]
Interpretation: Partial-report advantage \(A_{PR}\) measures how much more information is available when attention is cued to a subset of the sensory trace than when the whole array must be reported.
These simple formulations help express the core logic of sensory memory: brief persistence, rapid loss, modality-specific timing, selective stabilization, and report limitations.
Sensory memory and attention
Sensory memory captures far more information than can be processed consciously. The sensory environment contains enormous amounts of visual, auditory, and tactile information, most of which cannot be stabilized or interpreted in depth at the same time.
Selective attention therefore plays a decisive role in determining which sensory signals move beyond early registers into later stages of cognitive processing. Attention acts less like a passive observer and more like a selective gate. It helps determine which traces are enhanced, which are ignored, and which fade away before becoming available for report or active reasoning.
This is one reason sensory memory is so closely tied to attention. Early sensory traces provide the brief representational field from which attention can select, while attention helps determine which traces become usable input for working memory.
The relationship between sensory memory and attention also clarifies why early availability does not equal conscious access. Information can be present briefly in a sensory register without being selected, named, remembered, or reported. Attention is one mechanism through which a fleeting trace becomes part of later cognition.
In experimental terms, this distinction helps explain why partial-report performance can reveal more information than whole report. The information may be briefly present, but the capacity to select and stabilize it is limited. Sensory memory provides the field; attention determines what survives.
Sensory memory in the architecture of cognition
Sensory memory occupies the earliest position in the broader architecture of cognition. Within a classic information-processing framework, the system can be described in layered form:
- Sensory memory briefly preserves raw perceptual input.
- Attention selects a small subset of that input.
- Working memory maintains and manipulates selected information.
- Long-term memory preserves knowledge and experience across time.
This layered architecture helps explain how the mind manages the enormous volume of information generated by the sensory environment. Rather than processing every stimulus equally, cognition relies on hierarchical filtering and selective stabilization. Sensory memory is therefore not just a preliminary curiosity. It is part of the architecture that makes bounded cognition workable.
The architecture also shows why sensory memory matters for cognitive load. If early sensory information is poorly organized, rapidly overwritten, masked, or presented faster than attention can select it, later systems may be overloaded. Conversely, good perceptual design can support attention by making the relevant trace easier to select before it fades.
Sensory memory therefore sits at the boundary between the physical world and cognitive control. It is where stimulation first becomes available for selection, but not yet for full conceptual manipulation.
Sensory memory and perceptual continuity
Sensory memory plays a critical role in making perceptual experience feel continuous. Visual perception must integrate information across rapid eye movements, known as saccades. Auditory perception must integrate sounds distributed over time. Tactile perception must integrate contact as the body moves and interacts with surfaces.
Without brief sensory persistence, the world would seem much more discontinuous than it does. Visual scenes would fragment after each eye movement. Speech would be harder to assemble into meaningful phrases. Touch would be less able to integrate texture, pressure, and motion into stable object perception.
This means sensory memory is closely tied to perception. It provides the short temporal persistence that allows fragmented sensory input to become part of a more stable perceptual world.
Perceptual continuity does not require that all sensory information be preserved. It requires enough persistence for the system to bridge gaps, compare recent input with new input, and select what matters. Sensory memory helps provide that bridge.
In this way, sensory memory is not merely a memory system. It is also a condition of perceptual stability.
Masking, report limits, and access
Masking experiments are important for sensory-memory research because they show how fragile early sensory traces can be. A visual mask presented shortly after a stimulus can disrupt the availability of the original trace, reducing report accuracy even when the initial stimulus was present long enough to be registered.
Masking helps distinguish between several stages of processing. A stimulus may reach early sensory registration but fail to remain available for selection. It may be available briefly but overwritten before report. It may be selected but not stabilized. It may also be retained in a form that is perceptual rather than verbal or conceptual.
This matters because report is not a direct measure of sensory availability. Whole-report failure may reflect limits in attention, verbal report, working memory, or response production, not only absence of the sensory trace. Partial-report methods were powerful precisely because they separated early availability from later report capacity.
Researchers therefore need to interpret sensory-memory data carefully. A low report score does not always mean nothing was encoded. A high trace estimate does not always mean conscious access. Sensory memory occupies a narrow window between physical stimulation and later cognition, and experimental tasks must be designed to identify which part of that window is being measured.
This methodological caution remains important for contemporary debates about perception, consciousness, visual richness, and the relationship between availability and report.
Neuroscience and sensory persistence
Contemporary neuroscience suggests that sensory memory should not be understood as a single centralized memory container. Instead, sensory persistence appears to involve short-lived activity within modality-specific sensory and perceptual systems. Visual traces are linked to visual processing networks, auditory persistence to auditory cortical and temporal processing, and haptic persistence to somatosensory systems.
This distributed view fits the functional structure of sensory memory. The earliest memory traces preserve information in formats closely tied to the sensory systems that produced them. Iconic memory is visual. Echoic memory is auditory. Haptic memory is tactile. The traces are not yet abstract, semantic, or fully manipulable. They are still close to perception.
Neuroscientific work also helps explain why sensory memory is both fragile and useful. Short-lived activity can preserve information long enough for selection without requiring durable storage. This makes sensory memory efficient: most information fades quickly, while selected information can be routed into working memory, recognition, or action.
The neuroscience of sensory memory also matters for clinical and developmental research. Differences in sensory persistence, auditory integration, masking sensitivity, aging, sensory impairment, or attentional selection may affect speech processing, reading, interface use, warning detection, and everyday perception.
In this sense, sensory memory links cognitive psychology to neuroscience, human factors, accessibility, and applied perception.
Sensory memory, interface design, and human factors
Sensory memory has practical importance for interface design and human factors. Displays, alarms, dashboards, notifications, signs, haptic signals, and warning systems all rely on brief sensory availability. If critical information disappears before users can attend to it, the system may fail even if the information was technically presented.
Designers should therefore consider:
- how long information remains visible, audible, or tactile;
- whether users have time to select the relevant cue;
- whether masking or interruption disrupts perception;
- whether multiple cues compete for attention;
- whether a signal should persist, repeat, or be logged;
- whether users differ in sensory ability, fatigue, age, stress, or accessibility needs;
- whether auditory, visual, and haptic channels should reinforce each other.
Sensory-memory constraints are especially important in high-stakes systems: medical devices, aircraft displays, transportation alerts, industrial control rooms, emergency warnings, accessibility interfaces, and public safety systems. A warning that is too brief, too ambiguous, poorly timed, or easily masked may not support action.
Good design respects the difference between presentation and perception. A system has not communicated simply because it displayed a signal. The signal must remain available long enough, clearly enough, and accessibly enough for attention to select it and cognition to use it.
Sensory memory and artificial intelligence systems
Artificial intelligence systems do not have sensory memory in the human psychological sense, but the comparison is useful. AI perception systems often include short-term buffers, input windows, temporal smoothing, frame histories, audio segments, sensor fusion, and attention mechanisms that allow recent data to influence current processing.
The analogy should not be overstated. Human sensory memory is embodied, biological, modality-specific, and tied to perceptual experience. AI buffers are engineered structures. Still, both human and artificial systems face a similar computational problem: incoming signals change rapidly, and the system must preserve recent input long enough to detect patterns, select relevant information, and support action.
This comparison is especially useful in human-AI interaction. Users often interact with systems that present transient alerts, generated text, visual signals, audio prompts, or haptic feedback. If the system assumes that users can perceive and retain fleeting signals without support, it may overload or mislead them.
AI systems can support human sensory and attentional limits by:
- making alerts persistent or recoverable;
- avoiding unnecessary masking or interruption;
- using multimodal redundancy where appropriate;
- logging critical information after brief presentation;
- adapting timing to task context and user need;
- supporting accessibility for users with sensory differences;
- distinguishing urgent sensory alerts from routine notifications.
Sensory memory therefore matters not only as a psychological topic, but also as a design constraint for intelligent systems that interact with human perception.
Sensory memory in contemporary cognitive science
Current research on sensory memory draws from experimental psychology, neuroscience, psychophysics, attention research, perception science, human factors, and computational modeling. Reviews of early visual memory suggest that iconic memory is not a single simple store but may involve multiple components with different temporal and capacity properties.
Auditory research likewise distinguishes short and longer forms of auditory storage, emphasizing that echoic memory is not merely a visual-memory analogue in another modality. Auditory perception has its own temporal structure, and the persistence required for speech and sound processing differs from the persistence required for visual arrays.
Haptic research expands the field by showing that tactile sensory memory deserves more systematic study. Touch is not simply a weaker version of vision or audition. It is an active, bodily, exploratory modality with its own timing and integration demands.
Neuroscientific accounts suggest that sensory persistence emerges from short-lived activity within sensory cortical systems rather than from a single centralized memory structure. Computational models, meanwhile, allow researchers to estimate decay rates, selection probabilities, masking effects, and modality-specific timing parameters.
Sensory memory remains important because it reveals a foundational fact about cognition: before the mind can remember, reason, decide, or act, the world must persist long enough to be selected.
R code for sensory-memory data
The following R workflow illustrates analyses relevant to sensory-memory experiments, including cue delay, modality, partial report, masking, trace strength, attentional priority, report accuracy, working-memory transfer, response time, and modality-specific decay.
# Install packages if needed:
# pak::pak(c("tidyverse", "lme4", "lmerTest", "emmeans", "broom.mixed"))
library(tidyverse)
library(lme4)
library(lmerTest)
library(emmeans)
library(broom.mixed)
# Expected columns:
# participant, modality, condition, trial, stimulus_id,
# cue_delay_ms, stimulus_duration_ms, array_size, cue_validity,
# mask_present, trace_strength, salience, attentional_priority,
# report_score, correct, selection_probability, wm_transfer,
# rt_ms, confidence, perceptual_continuity
dat <- read_csv("sensory_memory_trials.csv") %>%
mutate(
participant = factor(participant),
modality = factor(modality),
condition = factor(condition),
stimulus_id = factor(stimulus_id),
correct = as.integer(correct),
mask_present = as.integer(mask_present),
wm_transfer = as.integer(wm_transfer),
log_rt = log(rt_ms)
)
# -----------------------------
# 1. Accuracy by cue delay
# -----------------------------
delay_summary <- dat %>%
group_by(modality, cue_delay_ms) %>%
summarise(
n_trials = n(),
accuracy = mean(correct, na.rm = TRUE),
mean_report = mean(report_score, na.rm = TRUE),
mean_trace = mean(trace_strength, na.rm = TRUE),
mean_selection = mean(selection_probability, na.rm = TRUE),
wm_transfer_rate = mean(wm_transfer, na.rm = TRUE),
.groups = "drop"
)
ggplot(delay_summary, aes(x = cue_delay_ms, y = accuracy, color = modality)) +
geom_point() +
geom_line() +
labs(
title = "Sensory trace accuracy across cue delay",
x = "Cue delay in milliseconds",
y = "Correct report rate"
) +
theme_minimal()
# -----------------------------
# 2. Correct-report model
# -----------------------------
correct_model <- glmer(
correct ~
cue_delay_ms * modality +
condition +
stimulus_duration_ms +
array_size +
cue_validity +
mask_present +
trace_strength +
salience +
attentional_priority +
(1 | participant) +
(1 | stimulus_id),
data = dat,
family = binomial(),
control = glmerControl(optimizer = "bobyqa")
)
summary(correct_model)
emmeans(correct_model, ~ modality, type = "response")
# -----------------------------
# 3. Report-score model
# -----------------------------
report_model <- lmer(
report_score ~
cue_delay_ms * modality +
condition +
array_size +
cue_validity +
mask_present +
trace_strength +
salience +
attentional_priority +
(1 | participant) +
(1 | stimulus_id),
data = dat,
REML = FALSE
)
summary(report_model)
# -----------------------------
# 4. Working-memory transfer model
# -----------------------------
transfer_model <- glmer(
wm_transfer ~
cue_delay_ms +
modality +
condition +
trace_strength +
correct +
cue_validity +
attentional_priority +
mask_present +
(1 | participant) +
(1 | stimulus_id),
data = dat,
family = binomial(),
control = glmerControl(optimizer = "bobyqa")
)
summary(transfer_model)
emmeans(transfer_model, ~ modality, type = "response")
# -----------------------------
# 5. Response-time model
# -----------------------------
rt_model <- lmer(
log_rt ~
cue_delay_ms * modality +
condition +
array_size +
correct +
mask_present +
attentional_priority +
confidence +
(1 | participant) +
(1 | stimulus_id),
data = dat,
REML = FALSE
)
summary(rt_model)
# -----------------------------
# 6. Perceptual-continuity model
# -----------------------------
continuity_model <- lmer(
perceptual_continuity ~
modality +
condition +
trace_strength +
wm_transfer +
mask_present +
cue_delay_ms +
confidence +
(1 | participant) +
(1 | stimulus_id),
data = dat,
REML = FALSE
)
summary(continuity_model)
# -----------------------------
# 7. Exponential decay estimates by modality
# -----------------------------
decay_estimates <- delay_summary %>%
filter(accuracy > 0) %>%
group_by(modality) %>%
group_modify(~{
out <- tryCatch(
{
fit <- nls(
accuracy ~ a * exp(-lambda * cue_delay_ms),
data = .x,
start = list(a = 0.9, lambda = 0.001),
control = nls.control(maxiter = 200)
)
tibble(a = coef(fit)[["a"]], lambda = coef(fit)[["lambda"]])
},
error = function(e) tibble(a = NA_real_, lambda = NA_real_)
)
out
}) %>%
ungroup()
print(decay_estimates)This workflow can be adapted for visual partial-report experiments, auditory partial-report analogues, tactile change-detection studies, masking paradigms, multimodal warning systems, attention-cueing tasks, perceptual-continuity research, and human factors experiments. Researchers should model participant and stimulus effects whenever possible because sensory persistence varies across people, stimuli, modalities, cue timing, masking conditions, and task demands.
Python code for sensory-memory data
The Python examples below parallel the R workflow and are useful for cue-delay paradigms, partial-report experiments, modality comparisons, masking studies, and sensory-trace decay modeling.
import numpy as np
import pandas as pd
import statsmodels.formula.api as smf
import statsmodels.api as sm
import matplotlib.pyplot as plt
from scipy.optimize import curve_fit
# Expected columns:
# participant, modality, condition, trial, stimulus_id,
# cue_delay_ms, stimulus_duration_ms, array_size, cue_validity,
# mask_present, trace_strength, salience, attentional_priority,
# report_score, correct, selection_probability, wm_transfer,
# rt_ms, confidence, perceptual_continuity
df = pd.read_csv("sensory_memory_trials.csv")
categorical_cols = ["participant", "modality", "condition", "stimulus_id"]
for col in categorical_cols:
df[col] = df[col].astype("category")
df["correct"] = df["correct"].astype(int)
df["mask_present"] = df["mask_present"].astype(int)
df["wm_transfer"] = df["wm_transfer"].astype(int)
df["log_rt"] = np.log(df["rt_ms"])
# -----------------------------
# 1. Accuracy by cue delay
# -----------------------------
delay_summary = (
df.groupby(["modality", "cue_delay_ms"], observed=True)
.agg(
n_trials=("correct", "size"),
accuracy=("correct", "mean"),
mean_report=("report_score", "mean"),
mean_trace=("trace_strength", "mean"),
mean_selection=("selection_probability", "mean"),
wm_transfer_rate=("wm_transfer", "mean"),
)
.reset_index()
)
fig, ax = plt.subplots(figsize=(8, 5))
for modality, group in delay_summary.groupby("modality", observed=True):
ax.plot(group["cue_delay_ms"], group["accuracy"], marker="o", label=str(modality))
ax.set_xlabel("Cue delay in milliseconds")
ax.set_ylabel("Correct report rate")
ax.set_title("Sensory trace accuracy across cue delay")
ax.legend(title="Modality")
plt.tight_layout()
plt.show()
# -----------------------------
# 2. Correct-report model
# -----------------------------
correct_model = smf.glm(
"correct ~ cue_delay_ms * modality + condition "
"+ stimulus_duration_ms + array_size + cue_validity "
"+ mask_present + trace_strength + salience + attentional_priority",
data=df,
family=sm.families.Binomial(),
)
correct_result = correct_model.fit(
cov_type="cluster",
cov_kwds={"groups": df["participant"]},
)
print(correct_result.summary())
# -----------------------------
# 3. Report-score model
# -----------------------------
report_model = smf.ols(
"report_score ~ cue_delay_ms * modality + condition "
"+ array_size + cue_validity + mask_present + trace_strength "
"+ salience + attentional_priority",
data=df,
)
report_result = report_model.fit(
cov_type="cluster",
cov_kwds={"groups": df["participant"]},
)
print(report_result.summary())
# -----------------------------
# 4. Working-memory transfer model
# -----------------------------
transfer_model = smf.glm(
"wm_transfer ~ cue_delay_ms + modality + condition "
"+ trace_strength + correct + cue_validity "
"+ attentional_priority + mask_present",
data=df,
family=sm.families.Binomial(),
)
transfer_result = transfer_model.fit(
cov_type="cluster",
cov_kwds={"groups": df["participant"]},
)
print(transfer_result.summary())
# -----------------------------
# 5. Response-time model
# -----------------------------
rt_model = smf.ols(
"log_rt ~ cue_delay_ms * modality + condition "
"+ array_size + correct + mask_present "
"+ attentional_priority + confidence",
data=df,
)
rt_result = rt_model.fit(
cov_type="cluster",
cov_kwds={"groups": df["participant"]},
)
print(rt_result.summary())
# -----------------------------
# 6. Perceptual-continuity model
# -----------------------------
continuity_model = smf.ols(
"perceptual_continuity ~ modality + condition "
"+ trace_strength + wm_transfer + mask_present "
"+ cue_delay_ms + confidence",
data=df,
)
continuity_result = continuity_model.fit(
cov_type="cluster",
cov_kwds={"groups": df["participant"]},
)
print(continuity_result.summary())
# -----------------------------
# 7. Exponential decay fit by modality
# -----------------------------
def decay_fn(delay, a, lamb):
return a * np.exp(-lamb * delay)
decay_rows = []
for modality, group in delay_summary.groupby("modality", observed=True):
fit_df = group[group["accuracy"] > 0].copy()
try:
params, _ = curve_fit(
decay_fn,
fit_df["cue_delay_ms"].values.astype(float),
fit_df["accuracy"].values.astype(float),
p0=[0.9, 0.001],
maxfev=10000,
)
decay_rows.append({"modality": modality, "a": params[0], "lambda": params[1]})
except Exception as exc:
decay_rows.append({"modality": modality, "a": np.nan, "lambda": np.nan, "error": str(exc)})
decay_estimates = pd.DataFrame(decay_rows)
print(decay_estimates)
# -----------------------------
# 8. Export summaries
# -----------------------------
delay_summary.to_csv("sensory_memory_delay_summary.csv", index=False)
decay_estimates.to_csv("sensory_memory_decay_estimates.csv", index=False)The Python workflow is intentionally transparent and extensible. It can be expanded with Bayesian decay models, hierarchical psychometric functions, drift-diffusion models for report timing, masking-effect models, eye-tracking integration, auditory temporal-window estimation, haptic change-detection models, multimodal cueing studies, and interface experiments for transient visual, auditory, or tactile signals.
GitHub Repository
The companion repository provides reusable code and research scaffolding for studying sensory memory in cognitive psychology, including workflows for iconic memory, echoic memory, haptic memory, cue delay, partial report, masking, trace decay, attentional priority, selection probability, report accuracy, working-memory transfer, response time, confidence, and perceptual continuity.
Complete Code Repository
Access the full companion repository for this article, including reproducible analysis materials and multi-language code workflows for sensory-memory research.
Applications of sensory-memory research
Sensory-memory research has practical importance in several areas. It helps clarify how perceptual systems stabilize rapidly changing information, how attention selects from brief sensory traces, and how interface design can support or disrupt early perceptual processing. It also informs work in auditory comprehension, visual display design, haptic interface design, human factors, safety systems, accessibility, and computational models of early perception.
In visual design, sensory-memory research helps explain why briefly flashed information, rapidly disappearing alerts, or cluttered displays may fail even when users technically “saw” the signal. In auditory design, it helps explain why warnings, speech, and environmental sounds need appropriate timing and repetition. In haptic design, it helps explain why tactile signals must be strong, well-timed, and interpretable across bodily movement.
In education and accessibility, sensory-memory research supports better design for people with visual, auditory, tactile, attentional, or processing differences. Users do not all have the same sensory persistence, attentional control, or working-memory capacity. Interfaces and learning environments should not assume identical perceptual timing across all people.
More broadly, sensory memory matters because it helps explain how cognition deals with the sheer flow of stimulation arriving from the world. Before information can be reasoned about, remembered, or acted upon, it must first persist long enough to be selected. Sensory memory is one of the mechanisms that makes that persistence possible.
Conclusion
Sensory memory is the earliest stage of cognitive processing in which raw sensory input briefly persists after stimulation has ended. By preserving recent visual, auditory, and tactile information for a short interval, it allows attention and perception to operate on a more stable field of input rather than on vanishing fragments.
Cognitive psychology shows that this persistence is brief, modality-specific, and deeply important. Sensory memory is not a durable knowledge system, but it is one of the first mechanisms through which the mind transforms stimulation into structured experience.
The central lesson is that perception requires time. Even the earliest stages of cognition depend on short-lived traces that bridge the gap between the physical signal and the selected information that later systems can use. Without sensory memory, perceptual continuity would be far harder to achieve, and later cognitive systems would have far less to work with.
Related articles
- Cognitive Psychology
- Perception in Cognitive Psychology
- Attention in Cognitive Psychology
- Working Memory in Cognitive Psychology
- Memory in Cognitive Psychology
- Cognitive Load and Information Processing
- Cognitive Learning Processes
- Human-Computer Interaction in Cognitive Psychology
- Artificial Intelligence Systems
Further reading
- American Psychological Association (n.d.) Sensory memory. APA Dictionary of Psychology. Available at: https://dictionary.apa.org/sensory-memory.
- American Psychological Association (n.d.) Iconic memory. APA Dictionary of Psychology. Available at: https://dictionary.apa.org/iconic-memory.
- American Psychological Association (n.d.) Echoic memory. APA Dictionary of Psychology. Available at: https://dictionary.apa.org/echoic-memory.
- Bradley, C. and Pearson, J. (2012) ‘The sensory components of high-capacity iconic memory and visual working memory’, Frontiers in Psychology, 3, 355. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC3457081/.
- Camina, E. and Güell, F. (2017) ‘The neuroanatomical, neurophysiological and psychological basis of memory: Current models and their origins’, Frontiers in Pharmacology, 8, 438. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC5491610/.
- Coltheart, M. (1980) ‘Iconic memory and visible persistence’, Perception & Psychophysics, 27(3), pp. 183–228. PubMed record available at: https://pubmed.ncbi.nlm.nih.gov/6992093/.
- Cowan, N. (1984) ‘On short and long auditory stores’, Psychological Bulletin, 96(2), pp. 341–370. PubMed record available at: https://pubmed.ncbi.nlm.nih.gov/6385047/.
- Darwin, C.J., Turvey, M.T. and Crowder, R.G. (1972) ‘An auditory analogue of the Sperling partial report procedure: Evidence for brief auditory storage’, Cognitive Psychology, 3(2), pp. 255–267. Available at: https://www.sciencedirect.com/science/article/abs/pii/0010028572900072.
- Michaelian, K. and Sutton, J. (2017) ‘Memory’, Stanford Encyclopedia of Philosophy. Available at: https://plato.stanford.edu/entries/memory/.
- Neisser, U. (1967) Cognitive Psychology. New York: Appleton-Century-Crofts.
- Sperling, G. (1960) ‘The information available in brief visual presentations’, Psychological Monographs: General and Applied, 74(11), pp. 1–29. Available at: https://sites.socsci.uci.edu/~whipl/staff/sperling/PDFs/Sperling_PsychMonogr_1960.pdf.
- Sugiyama, S. et al. (2020) ‘Assessment of haptic memory using somatosensory change-related cortical responses’, Psychophysiology. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC7643370/.
References
- American Psychological Association (n.d.) Sensory memory. APA Dictionary of Psychology. Available at: https://dictionary.apa.org/sensory-memory.
- Bradley, C. and Pearson, J. (2012) ‘The sensory components of high-capacity iconic memory and visual working memory’, Frontiers in Psychology, 3, 355. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC3457081/.
- Camina, E. and Güell, F. (2017) ‘The neuroanatomical, neurophysiological and psychological basis of memory: Current models and their origins’, Frontiers in Pharmacology, 8, 438. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC5491610/.
- Coltheart, M. (1980) ‘Iconic memory and visible persistence’, Perception & Psychophysics, 27(3), pp. 183–228. PubMed record available at: https://pubmed.ncbi.nlm.nih.gov/6992093/.
- Cowan, N. (1984) ‘On short and long auditory stores’, Psychological Bulletin, 96(2), pp. 341–370. PubMed record available at: https://pubmed.ncbi.nlm.nih.gov/6385047/.
- Darwin, C.J., Turvey, M.T. and Crowder, R.G. (1972) ‘An auditory analogue of the Sperling partial report procedure: Evidence for brief auditory storage’, Cognitive Psychology, 3(2), pp. 255–267. Available at: https://www.sciencedirect.com/science/article/abs/pii/0010028572900072.
- Michaelian, K. and Sutton, J. (2017) ‘Memory’, Stanford Encyclopedia of Philosophy. Available at: https://plato.stanford.edu/entries/memory/.
- Neisser, U. (1967) Cognitive Psychology. New York: Appleton-Century-Crofts.
- Sperling, G. (1960) ‘The information available in brief visual presentations’, Psychological Monographs: General and Applied, 74(11), pp. 1–29. Available at: https://sites.socsci.uci.edu/~whipl/staff/sperling/PDFs/Sperling_PsychMonogr_1960.pdf.
- Sugiyama, S. et al. (2020) ‘Assessment of haptic memory using somatosensory change-related cortical responses’, Psychophysiology. Available at: https://pmc.ncbi.nlm.nih.gov/articles/PMC7643370/.